広島大学 大学院 先進理工系科学研究科 先進セルロース材料共同研究講座
セルロース・複合材料の分析評価SERVICE&PRODUC
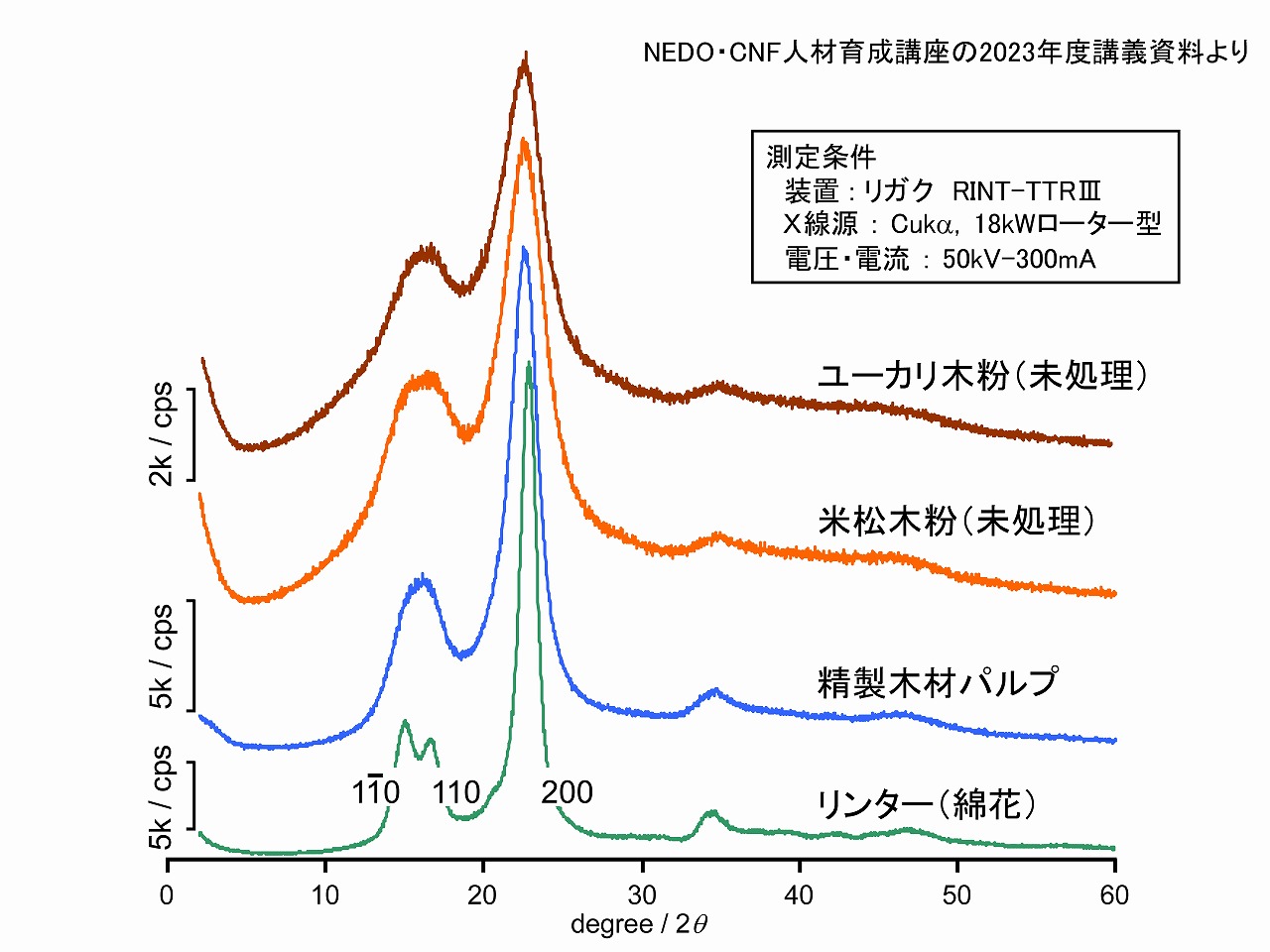
木材中の主要成分は、セルロース、ヘミセルロース、リグニンです。この中で、セルロースはセルロース分子が規則正しく集合した構造を持つ、「セルロースミクロフィブリル」です。この規則正しい集合から結晶性を示します。セルロースを原料等として取り扱う場合、セルロースの結晶性は、場合によりとても重要となります。
木材からパルプを製造したり、木粉を樹脂と複合化する場合には、一般的には、セルロースはダメージを受けて結晶性が低下していきます。結晶性が低下したセルロース原料を複合材料等に利用すると、期待した補強効果が発揮されない場合もあります。
上図には、一般的なセルロース原料として、木材、パルプ、コットンリンター(綿花/精製済み)のエックス線回折パターンを示しました。代表的な回折ピークとして、低角度から3ピーク(11-0、110、200)があります。ピークに付けている3種の数値は、結晶構造の点群(数学的形状表記法)で示されるもので、日本語では、11-0は、イチ・イチ・バー、ゼロ、200は、ニー・ゼロ・ゼロと読みます。結晶の立体の位置なので、110面(イチ・イチ・ゼロ・めん)等と呼ばれる場合が一般的です
コットンリンターとパルプや木材(木粉)は15度付近の形が異なっていますが、これは、本来はコットンリンターが示すように2つのピークがあるところが、パルプや木材では結晶サイズが小さいためにブロードになって1つのように見えているだけです。
このようなエックス線回折ピークが鋭い(シャープ)な場合は、結晶性が高く、なだらか(ブロード)の場合は、結晶性が低いと考えることができます。上図では、左側の軸に5k/cps等と記載していますが、これは回折したエックス線を検出器がカウントした値です。単位のcpsは、1秒あたりの検出器のカウント数の意味です。この値は、エックス線回折装置により異なり、サンプルの調製方法によっても変わるため、ピークの高さを、他のデータと比較しても意味がありません。回折パターンで比較等するのが一般的です。また、ヘミセルロースやリグニンは非晶質(アモルファス)な物質であるため、回折ピークは出てきませんが、20度付近になだらかな山(ハローパターン)として現れるため、木材等の評価でのエックス線回折のパターンには、セルロース以外に非晶質のヘミセルロースやリグニンも重なって現れていることに注意が必要です。
※上図のエックス線回折を測定した装置は、粉末エックス線回折装置(広角エックス線回折装置)です。エックス線回折装置には、単結晶エックス線回折装置、薄膜エックス線回折装置、測定する領域により、広角エックス線回折装置、小角エックス線回折装置などがあります。
結晶と非晶質のエックス線回折評価
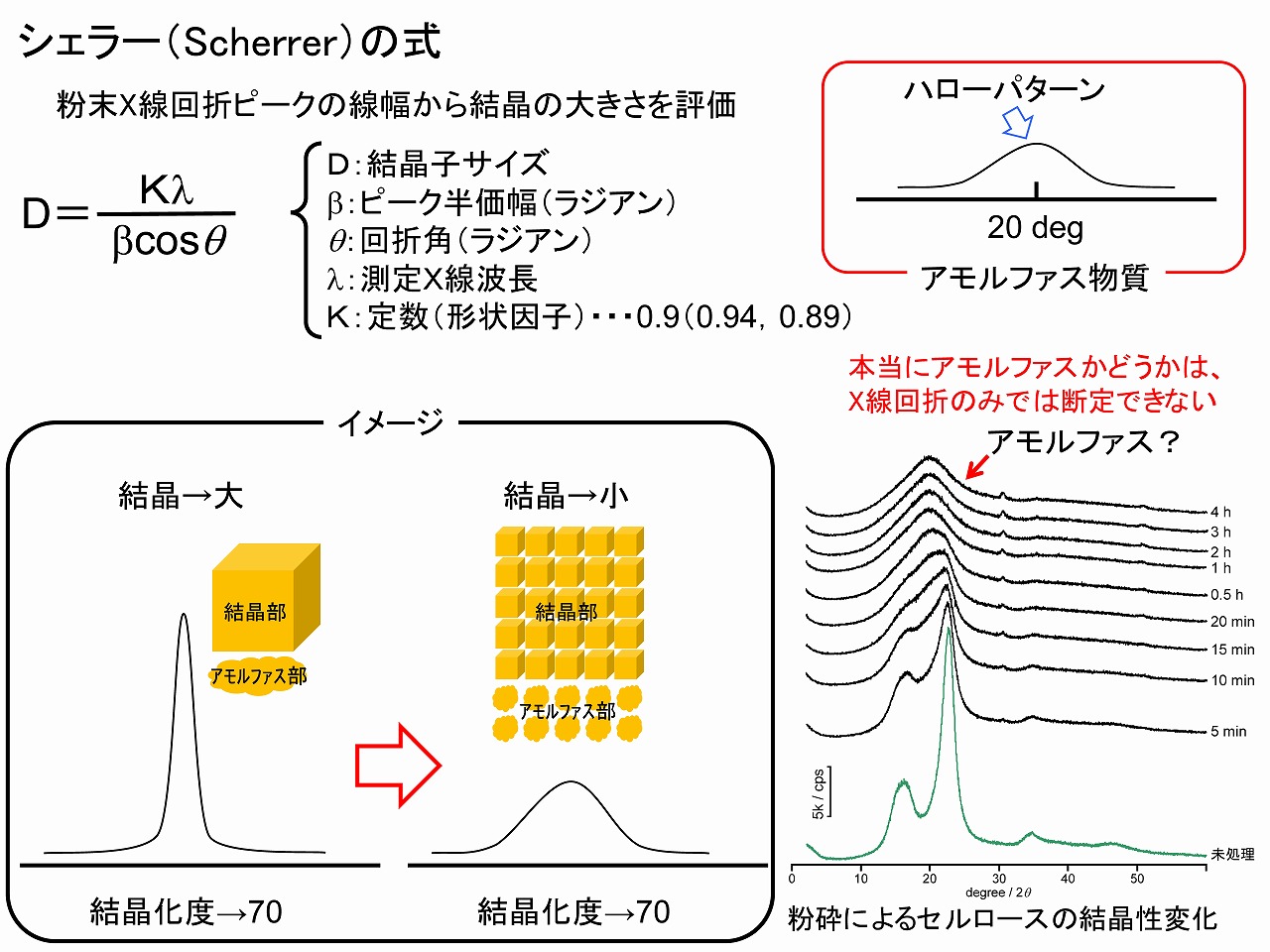
上の項目「セルロースの結晶性(エックス線回折測定)」でも記載したように、ヘミセルロースやリグニンの様な非晶質(アモルファス)な物質は、一般的なエックス線回折ではハローパターンを示します。セルロースの結晶の本体は、セルロースミクロフィブリルですが、そのサイズ(幅/直径)は3nm程度でとても微細です。エックス線回折ピークの鋭さは、結晶のサイズに影響されます。
結晶のサイズとピークの鋭さは、「シェラー(Scherrer)の式」で表すことができます。結晶のサイズとピークの鋭さ(上図で、ピーク半価幅)は、反比例の関係にあります。ピーク半価幅とは、ピークの高さの半分の位置の幅のことで、ピーク半価幅が小さいと言うことは、鋭いピークであることを意味します。このことは、結晶性が高い物質でも、その結晶サイズが小さいと、エックス線回折ではブロードに見えてしまうことを示しています。つまり、結晶性(結晶化度)は、物質の結晶サイズが小さい場合は、高結晶性なのか非晶質(アモルファス)なのか、判断できないことになります。
セルロースは本質的に結晶サイズが小さいため、処理プロセスによっては、さらに結晶サイズが小さくなる場合も考えられ、結晶性をキッチリ評価するためには、固体NMR等での評価方法も併用することも大切です。
結晶性のセルロース以外に非晶質のヘミセルロースやリグニンが含まれた材料、例えば、木粉等をエックス線回折で評価する場合、別途求めたヘミセルロース等の成分割合をハローパターンとして回折パターンから、差し引いてセルロースの結晶性を算出する方法やセルロース由来ピークをピーク分離によって算出する方法はありますが、誤差が大きくなる場合や計算している人の意思が入り、結果が正しく出ない場合もあります。そのため、同一系内での評価であれば、様々な回折を含んだ測定そのままの回折ピークを用いて議論する方が無難な場合も多いです。
※「シェラー(Scherrer)の式」:スイスの物理学者であった、パウル・シェラー(Paul Scherrer)が1918年に示した、粉末エックス線回折パターンから結晶の大きさを評価する計算式。
文献:P. Scherrer, Göttinger Nachrichten Gesell., Vol. 2, 1918, p 98.
エックス線回折によるセルロースの結晶化度算出法1
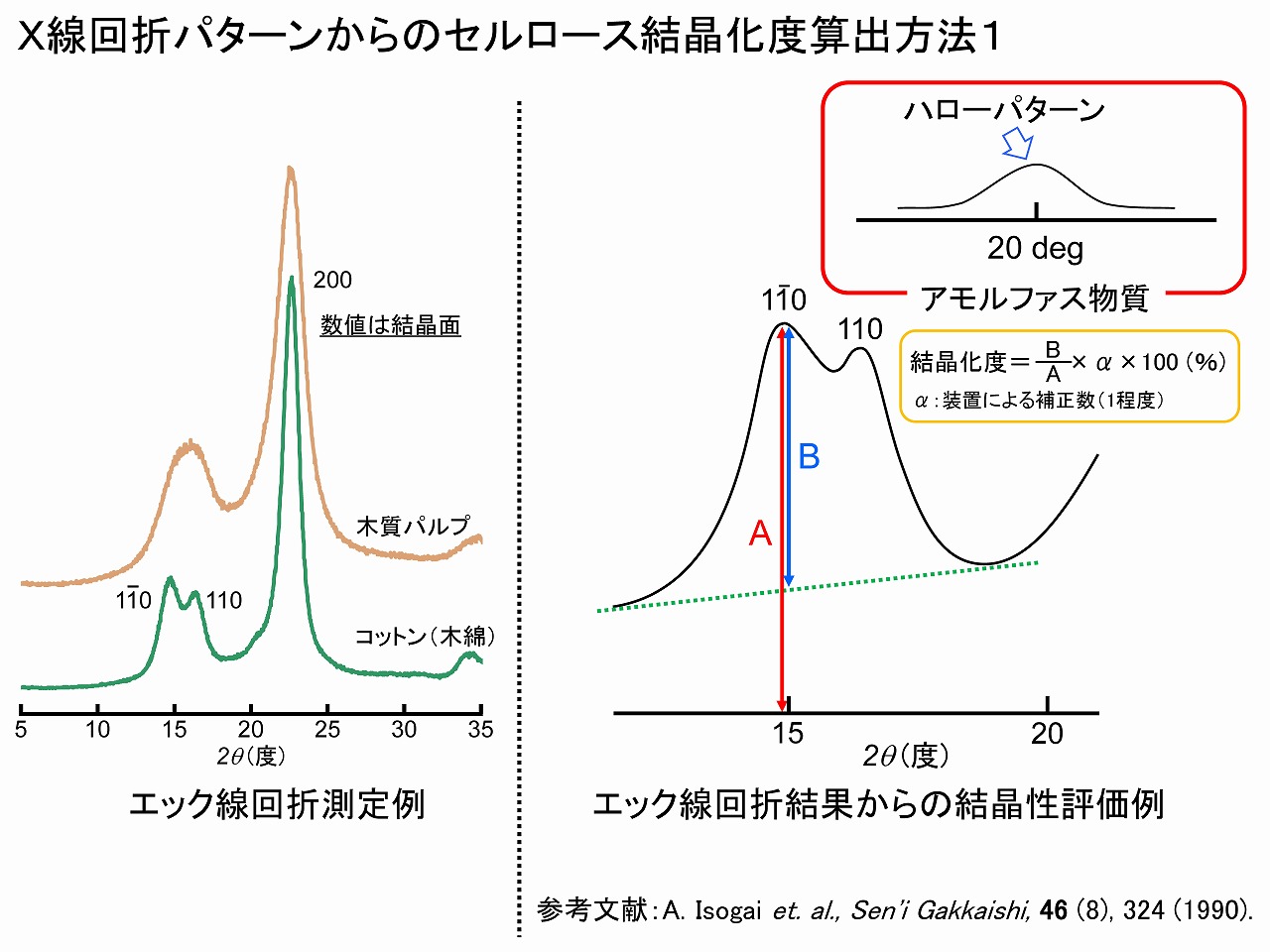
セルロースの結晶化度を算出する場合の注意点は、前項に記載しましたが、ここでは、実際の計算方法の事例を紹介します。上の「セルロースの結晶性(エックス線回折測定)」の項で示したように、セルロース原料のエックス線回折パターンは、由来によって異なっています。
算出方法1の図では、我々が用いている、東京大学・磯貝明教授の文献による方法を示しています。コットン(綿花)由来のセルロースでは。15度付近には、2つのピークが観察されます。この算出方法では、低角度の方の110面(イチ・イチ・ゼロ・めん)のピークを使って、左の方に示した計算式で算出します。ここの装置補正数は、我々は複数台のエックス線回折装置を使いましたが、「1」としました。その根拠は、例えば、結晶性の高いコットンリンター等のエックス線回折パターンを測定し、計算式に導入して、結晶化度が95%になるように装置補正数を決めました。エックス線回折パターンは、装置への依存性も高いため、同型系内での相対比較であれば、装置補正数は「1」で問題無いです。100%セルロース結晶の物質は無いと考えられているため、本当に真の値は分かりません。そのため、他機関のデータでのセルロースの結晶化度の値と直接的には比較はできません。この方法では、110面(イチ・イチ・ゼロ・めん)のピークを使っていますが、木粉や木材由来パルプは、2つピークが観察されません。エックス線回折データは、そもそもデータが誤差をそれなりに含んでいるため、我々は、ペースライン(上図では緑の破線)から見て、最も高い場所を基準として計算しています。セルロースの結晶化度計算値は誤差も大きいので、算出基準は、そのデータを必要とする機関で決めた方法で良いと思います。
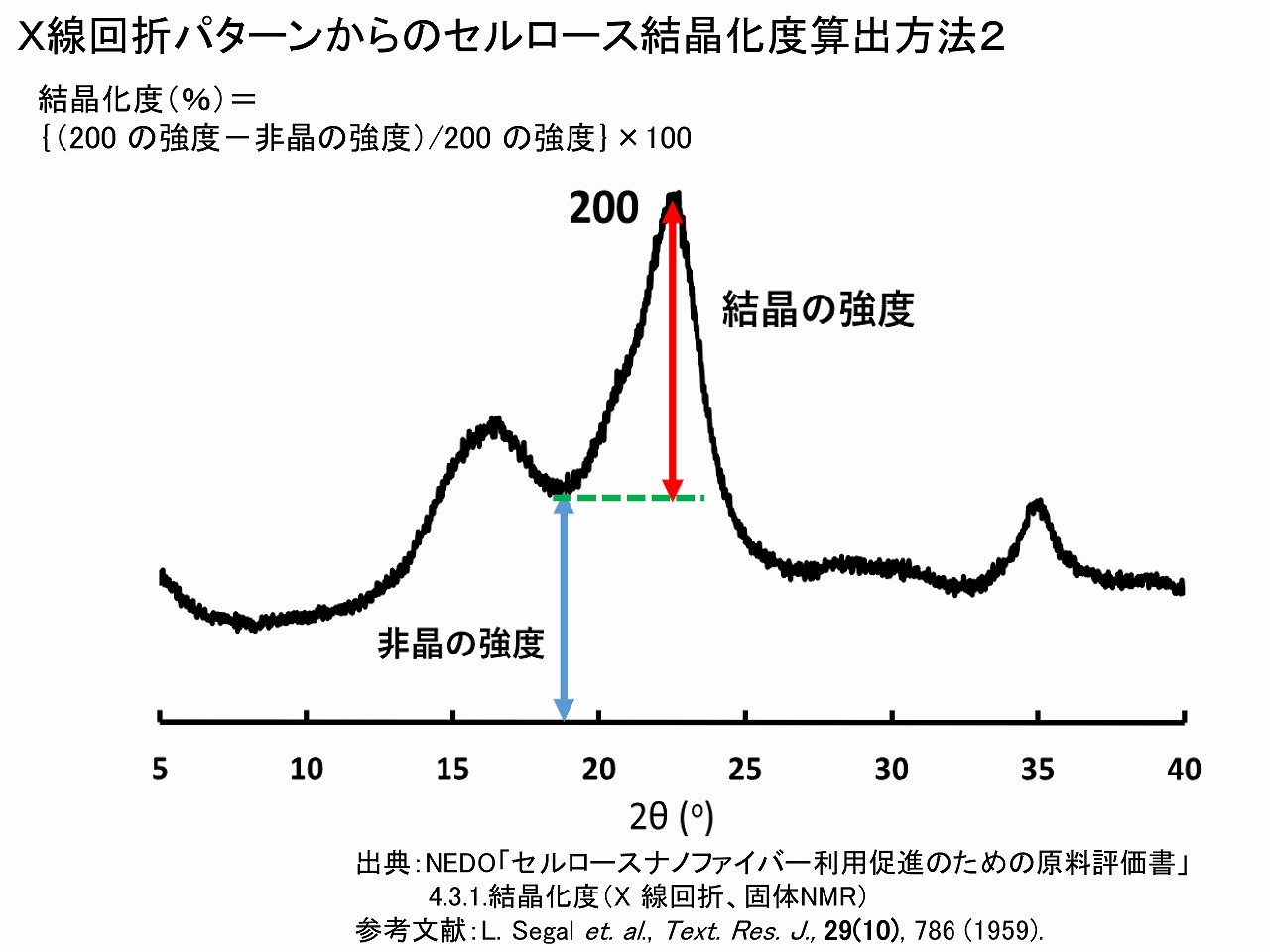
算出方法2の方法は、エックス線回折でのセルロースの結晶化度計算ではよく使われている方法です。提案者の名前を取って、「シーゲル法」とも呼ばれていますこの方法は、非晶質(アモルファス)の20度付近のハローパターンと部分的に重なっている200面の回折ピークを利用するため、結晶化度産出値の直線性が低いとされています。算出方法1の論文では、そのことが指摘されており、それを解決する方法として15度付近のピークを利用する方法が提案されています。
以上のように、セルロースの結晶化度は、装置、算出方法によって変わります。外部に発表・報告する際には、どのようにして値を求めたかは、併記・説明が必要です。
セルロースの固体NMR測定と結晶性
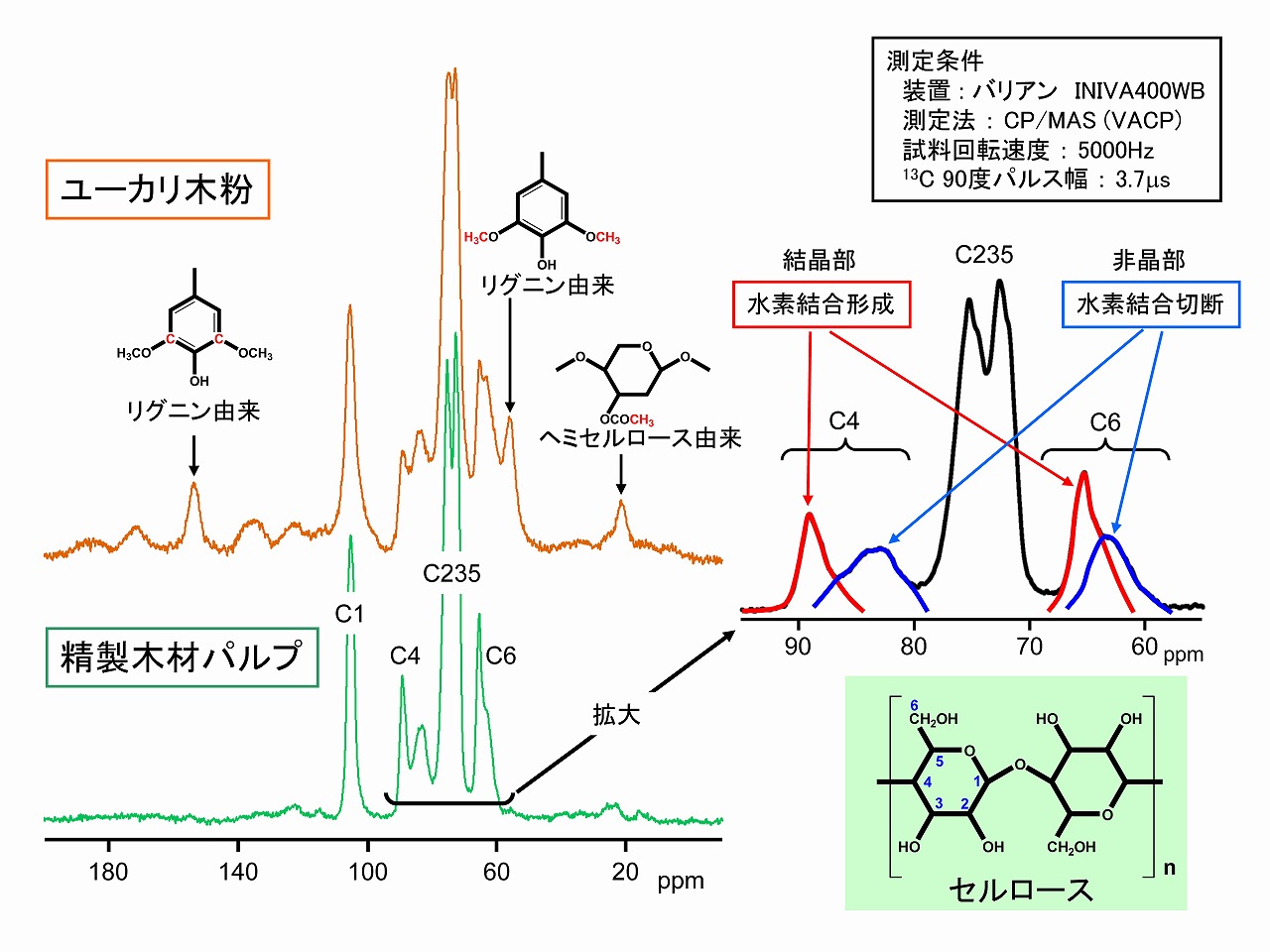
前の項に記載したように、エックス線回折のみでは、セルロースの結晶性を評価できない場合(結晶サイズがとても小さい場合)があるかもしれません。そのような場合には、固体NMR測定が有効です。とても汎用的な装置ではありませんが、セルロースの場合、結晶部分と非晶部分のピークが別の位置で観測されます。C4位とC6位です。この番号は炭素原子の順番を示すもので、分子構造から国際的基準に従って、1番(C1)、2番(C2)・・・と続きます。このC4位とC6位は、一般的にはセルロースの結晶と非晶を反映するとされていますが、セルロースの学術的考えでは、結晶と言うより、水素結合が正しく形成されているところと、乱れているところ(セルロースミクロフィブリルの表面なども)に由来するもので、必ずしも、結晶・非晶と区別はできないと言われています。
しかしながら、C4位とC6位のピークから、もし結晶サイズがとても小さくても、上図で示した赤線で示すピーク(低磁場側のピーク)が大きければ、結晶部分が多いと言えます。もし、エックス線回折で低結晶性に見えても、固体NMRで解析すると、実は高結晶で結晶サイズが小さいセルロース素材だったということもあるかもしれません。セルロースの解析は簡単ではないので、気になるときは、様々な分析手法を組み合わせて、総合的に評価することが大切です。
※NMRのピークの説明で用いる、低磁場側-高磁場側、の表現は、NMRの原理から来るもので、スペクトル図の、左側は、低磁場側、右側は高磁場側です。横軸のppmの値が大きくなる左側が低磁場側なので、勘違いに注意が必要です。
セルロースの偏光顕微鏡観察
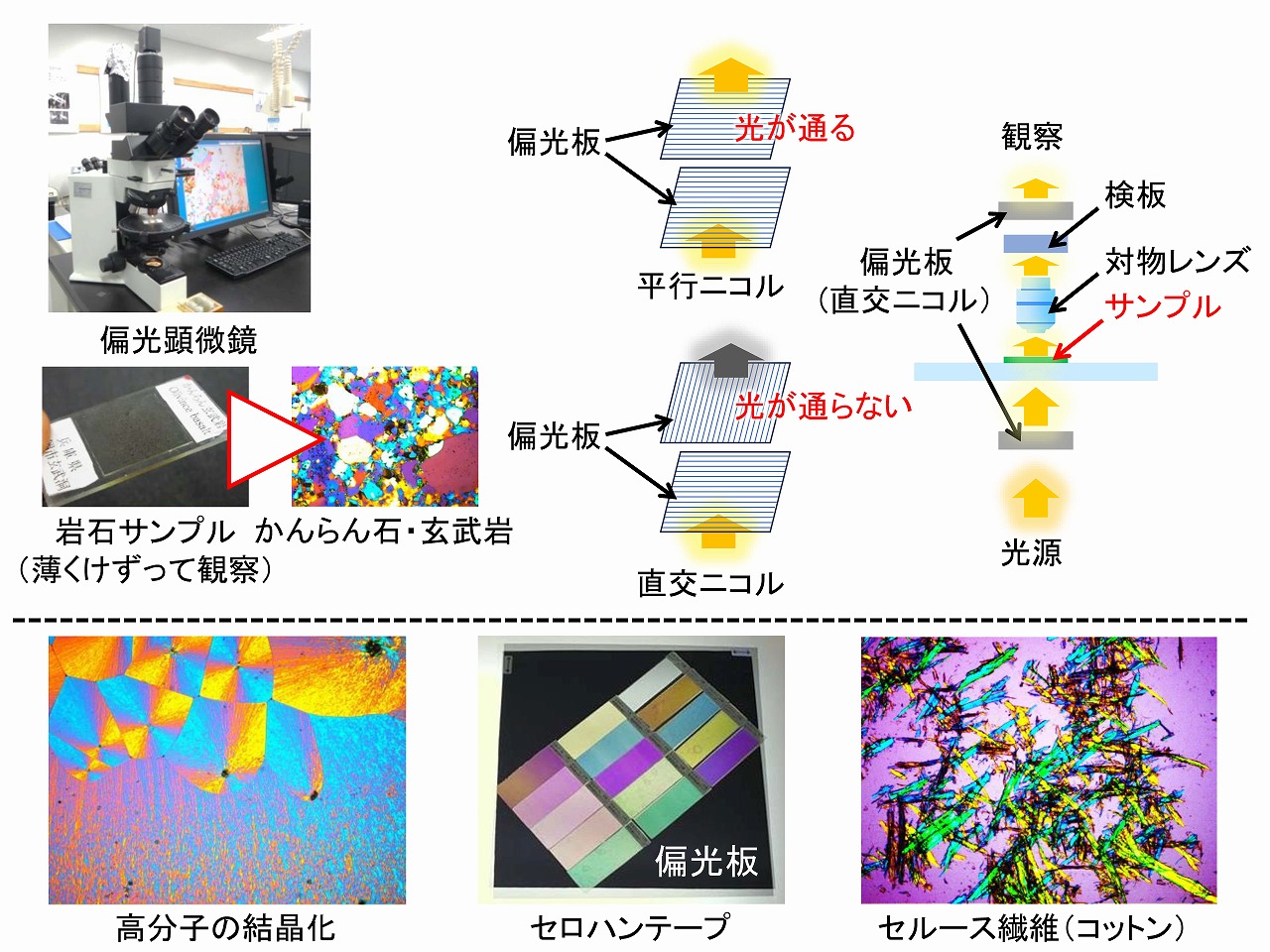
セルロースり基本単位は、セルロースミクロフィブリルです(参照:基礎解説/木材組織とセルロース/木質組織の基本構造)。このミクロフィブリルでは、セルロース分子が繊維軸方向に整列して、結晶構造を持っています。コットン繊維やパルプ繊維も、基本的には繊維軸方向にセルロースミクロフィブリルが並んでいます(コットンはらせん構造)。そのため、偏光性を示します。偏光性を示さない物質もありますが、偏光性物質を偏光顕微鏡で観察すると、特徴的に観察することができます。偏光顕微鏡は、岩石の構成成分の解析でよく使われている装置です。高分子系でも、分子配向などを調べるためにも使われています。
偏光顕微鏡は、光学顕微鏡(生物顕微鏡)に、2枚の偏光板が組み込まれています。検板(観察時の色を強調させる)も組み込まれる偏光顕微鏡もあります。偏光板は、一定方向に分子が整列した板で、サンプルを挟んで上下にセットされています。偏光板の分子方向を上下ともに平行にした観察は、平行ニコル(単ニコル、開放ニコル)と呼ばれ、90度交叉させた場合は、クロスニコル(直交ニコル、交叉ニコル)と呼ばれています。高分子素材の観察では、クロスニコル観察が多いです。通常の観察では、偏光板をクロスニコル状態にしてサンプルをステージにセットし、ステージを回転させて解析します。たまに、上部の偏光子を回転させて観察する人がいますが、基本は、クロスニコルでサンプルステージを回転させて調べます。検板を用いると、サンプルによっては白黒だったものが、カラフルな白になることも多いです。そのため、検板は鋭敏色板とも言われます。検板には幾つかの種類があり、サンプルや目的に応じて使い分けします。
ちなみに、ニコルとは、偏光を作り出す方解石のプリズムを発明(1825年)した、ウイリアム・ニコル(イギリス)さんから来ています。

上図は、特殊な方法で、コットン繊維から製造した特殊形状のセルロース粉体の偏光顕微鏡写真です。ここでは、クロスニコルで、鋭敏職板を挿入して観察しました。左方向に傾いた矩形状のセルロース粉体は赤色系、右方向に傾いたものは青系になっています。1つの矩形状セルロース粉体中では、それぞれ全体が同じ色になっています。偏光顕微鏡観察での色は、サンプルの厚みによっても変わってきますが、このサンプルは、とても薄いもので、向きによって色が決まっており、この矩形状セルロース粉体では、長軸方向にセルロース分子が配向していると考えることができます。もし、偏光顕微鏡で観察しても、原料セルロースでは明確に方向によって色が決まっているのに、処理後などでは、色があまり明確には現れず、サンプルを回転させても、常に白色に見える場合は、セルロース分子は特定方向に配向しておらず、様々な向きのセルロース分子がランダムに混在していると考えられます。
偏光顕微鏡によるセルロース繊維の評価
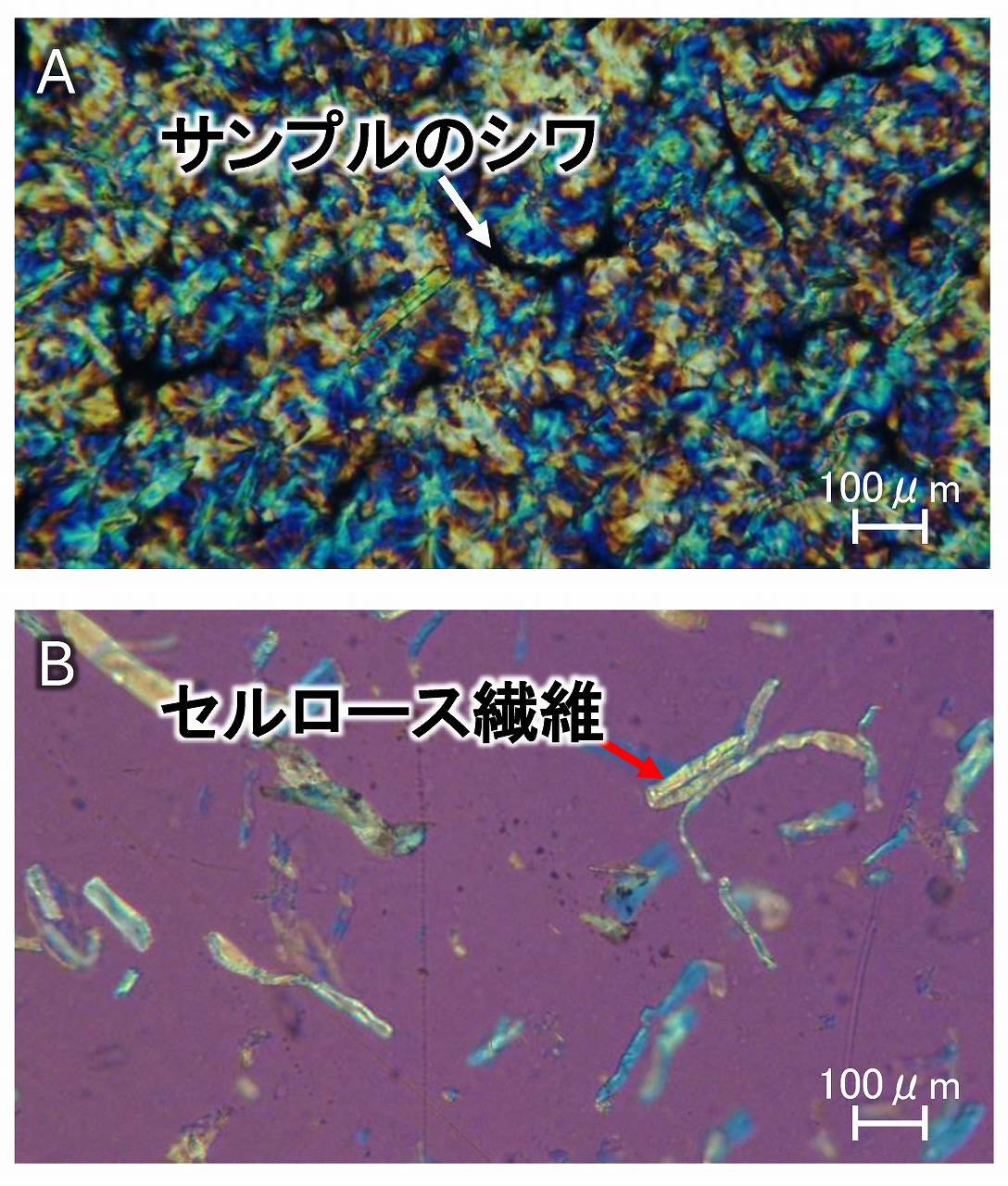
木粉やパルプ等のセルロース繊維を樹脂等に複合化した場合、樹脂中でのセルロース繊維の分散状態などは、複合材料の強度などの物性に影響するため、とても気になります。複合化するあいての樹脂としてポリプロピレン(PP)はよく研究・開発されていますが、PPが結晶性樹脂のため、偏光顕微鏡などで観察しようとすると、PPの結晶が邪魔する場合があります。上図のA(鋭敏色板使用)は、モデル的に木粉系セルロース繊維を、少量の約2wt%程度、溶融混練で複合化しただけの複合材料のシートの偏光顕微鏡画像ですが、シートをホットプレスで成形後にゆっくりと冷却したことでPPの球晶が成長して、セルロース繊維の観察が困難です。これまで、サンプルを顕微鏡用ホットステージなどにセットして、温度を180℃程度に向上させて、PPを溶融させて結晶を消して、観察する方法などがありますが(セルロースは溶融しないので、偏光顕微鏡ではセルロースのみが観察できます)、顕微鏡用ホットステージはとても高価で,手間もかかります。
上図BはAと同一サンプルですが、PPの結晶は見えず、セルロース繊維のみが観察されています。このサンプルは、ホットプレスでシート成形後にただちに冷水等で急冷(液体窒素を使わなくても、氷水で十分)したのみのサンプルです。急冷によりPPの結晶化は抑制されるため、偏光顕微鏡では何も球晶の模様などは見えてきません。見えるのは、セルロース繊維のみです。ホットプレスでシート化するため、射出成形等での成形体そのものでのセルロース繊維の状態とは異なってしまいますが、セルロース繊維の凝集体の有無などは確認できると思います。とても簡単な方法なので、PP複合材料中のセルロース繊維の簡易的状態観察に使える方法と思います。

偏光顕微鏡でのセルロース繊維の観察では、繊維軸等に対する色もセルロース繊維のダメージの評価にも使えます。セルロース繊維では、多くの場合で、繊維軸方向にセルロース分子が配向しています。配向が揃っていると、偏光顕微鏡で観察すると、同一の色に見えます。上図は偏光顕微鏡に鋭敏色板をセットしているため、ある角度では青、反対の角度では赤やオレンジに見えます。もし、色がキレイに揃わず、乱れている場合は、そのセルロース繊維(粉体)でのセルロース分子の配向が乱れている可能性があり、複合材料での繊維補強効果が低下する可能性があります。
上図は、日本製紙のKCフロック・W100G単体を、乾式でボールミル粉砕した場合のセルロース繊維(粉体)の偏光顕微鏡での観察像の変化を示しています。未処理(原料そのまま、未粉砕)では、セルロース繊維軸に沿って、キレイな色が出ています。しかし、90分粉砕後の粒子は特定の色が観察されず、セルロース分子の配向がとても乱れていることが分かります。粉砕後でもダメージをあまり受けていない部分では、色がしっかり出ていますが、とても小さい領域のみです。上図の左下の図では、未処理の場合には、微細な繊維でも全体的に色は単一で、セルロース分子はキレイに配向していることが分かります。
樹脂複合材料中でのセルロース繊維の偏光顕微鏡観察では、複合化や混練でセルロース繊維がダメージを受けていないかどうかの簡易評価にも偏光顕微鏡は使うことができると思います。セルロース繊維にダメージがあると、複合材料の物性は低下するかもしれません。安価な偏光顕微鏡は、それなりの性能でも10万円程度からあります。シート成形と偏光顕微鏡観察は、工場の現場での簡単な品質管理方法とし活用できるかもしれません。
※有名メーカーのホットプレスは高価です。ネット通販サイト等で、「ホットプレス あるいは 熱プレス」と入れて検索すると、ステージは10×5センチ程度ですが、10トンのホットプレスが、4~5万円程度で販売されています。性能と安全性は保証できませんが、我々の講座でも安価(安物)ホットステージを活用しています。購入は自己責任で!!!
簡易的蛍光顕微鏡観察
セルロースは蛍光染色されやすい物質で、セルロースを特異的に蛍光染色して、存在状態等をしらべる手法は、セルロース研究でもよく実施されています。当研究講座でも類似の研究を実施していますが、蛍光顕微鏡観察そのものについて、もっと簡単に、もっと低コストで実施できないか色々と試行錯誤しました。ここでは、簡易的ではあるものの、普通の安価な光学顕微鏡を用いて、とても手軽に実施できる方法について紹介しています。市販のセルロース繊維や紙などは、白く見せるために蛍光染色されているものがたくさんあります。ブラックライト(UVライト)を当てると、強く青白く光ります。
蛍光顕微鏡は材料からバイオ分野まで、昔から幅広く利用されている分析装置です。一般的な蛍光顕微鏡の基本は光学顕微鏡であるため、原理的にも電子顕微鏡のような分解能を得ることができません。しかし、近年開発された共焦点レーザー顕微鏡は大きく進歩しており、ナノレベルの分解能を発揮します。この技術は、「超高解像蛍光顕微鏡の開発」として2014年に開発に携わったエリック・ベッチグ(米国)、シュテファン・ヘル(ドイツ)、ウィリアム・モーナー(米国)の3氏がノーベル化学賞を受賞しています。しかしながら、共焦点レーザー顕微鏡はまだまだ高価で、簡単には導入できません。
従来からの蛍光顕微鏡は、分解能と言うよりは、特定の組織や物質を特異的に蛍光染色することで染め分けて、どこにどのように組織・物質が存在しているかを調べる目的で使われることも多いです。蛍光顕微鏡の光源は基本的には紫外線を発する必要があります。歩くからは、高圧水銀ランプが使われきました。高圧水銀ランプは、光量は大きいものの発熱し、ランプ寿命も長くありません。さらに、世界的に水銀の害が問題となり、日本政府主導で進められた国際連合環境計画「水銀に関する水俣条約」により、2021年から一般照明用の高圧水銀灯の製造・輸出・輸入が禁止となっています。最近のCM等で告知されているように、水銀を用いているため蛍光灯も2027年から製造・輸入が禁止となります。そのため、LEDタイプの照明器具等への切替が推奨されています。現在、蛍光顕微鏡用の水銀ランプは流通量が減少して値上がりしとています。
蛍光顕微鏡も、近年の製品は光源がLEDになったものが主流になっています。LEDは長寿命かつ低消費電力です。しかし、今までの蛍光顕微鏡の水銀ランプを簡単にLEDランプに交換することは、とても困難です。水銀ランプ型の蛍光顕微鏡をLED型の蛍光顕微鏡に買い換えたら良いのですが、蛍光顕微鏡には、ランプ以外にも専用フィルターなどが内蔵されており、低価格な顕微鏡ではありません。また、今まで蛍光顕微鏡は使っていなかったが、研究開発のテーマからの蛍光顕微鏡が必要となり新たな導入を検討することもあるかもしれません。
蛍光顕微鏡観察では、光をサンプルに照射して(励起光)、蛍光を発する部分・構造・組織を顕微鏡で観察します。励起光を変えることで、UV(紫外)、赤、緑、青で観察します。蛍光顕微鏡には、この切替機能が付いています。サンプルによりますが、UVでの蛍光は大きく、他の色(波長)よりは観察がし易い特徴があります。
少し前までは、紫外を発するLEDは出力が小さく、高出力のものは高価でした。しかし、近年は、365nmなどの低波長のLEDも普通になってきました。汚れ検出などで、大手通販サイトなでも、「365nm」等で検索すると懐中電灯タイプのものがたくさん出てきます。懐中電灯タイプではなく、照射専用型も販売されています。その目的は、最近、工作や手芸分野で流行している「UV硬化レジン」に使う大丈夫めです。ネール用としても使われています。ただ、紫外線なので皮膚に長時間当てることは避けなければなりません。特に、「眼」は注意です。安全メガネの利用も大切です。安価で十分な性能の紫外線カット安全メガネも市販されています。(例:アズワン・2-9048-04 紫外線用メガネ(オーバーメガネタイプ) SSUV-2060)

上図は、大手通販サイトで購入した365nmのUVランプです。価格も2~4千円程度です。USB電源に繋いで使用します使用します。消費電力は、5V-1A程度ですので、安価なUSB
電源で十分です。途中にON-OFFスイッチも付いています。
LEDランプ単体が入手できず、上図の懐中電灯タイプのものしかない場合は、上図の下の写真のようなフレキシブルクランプに保持してプレパラートに照射することも簡単です。LEDランプタイプタイプも、これらフレキシブルクランプに保持して照射しました。写真で懐中電灯型UVライトを保持しているものは、フレキシブルのマイクスタンドです。

上図は、格安の365nm LED UVランプをプレキシブルクランプに固定して、光学顕微鏡(生物顕微鏡)にセットしたプレパラートの横上部からUVを照射しているところです。プレパラートには、蛍光染色したセルロース系サンプルがセットしてあります。UV
LEDランプは、なるべくプレパラートに近い方が光量は大きくなります。あまり斜めから照射すると高倍率時に画像のピントが甘くなります。我々は、LED
UVランプ照射の場合は、UV直視にならないように、CCDラメラを顕微鏡に接続した観察を基本としています。必要に応じて、暗幕で全体を覆います。室内照明の影響低減と、UVの漏れでの皮膚照射を防止しています。
◇4K CCD(高性能型):(株)レイマー 顕微鏡用マルチインターフェース4KカメラFLOYD-4K
◇4k CCD(汎用型):HAYEAR社 4K 41MP HDMI/USB 3.0 デュアル インターフェース HY-6110
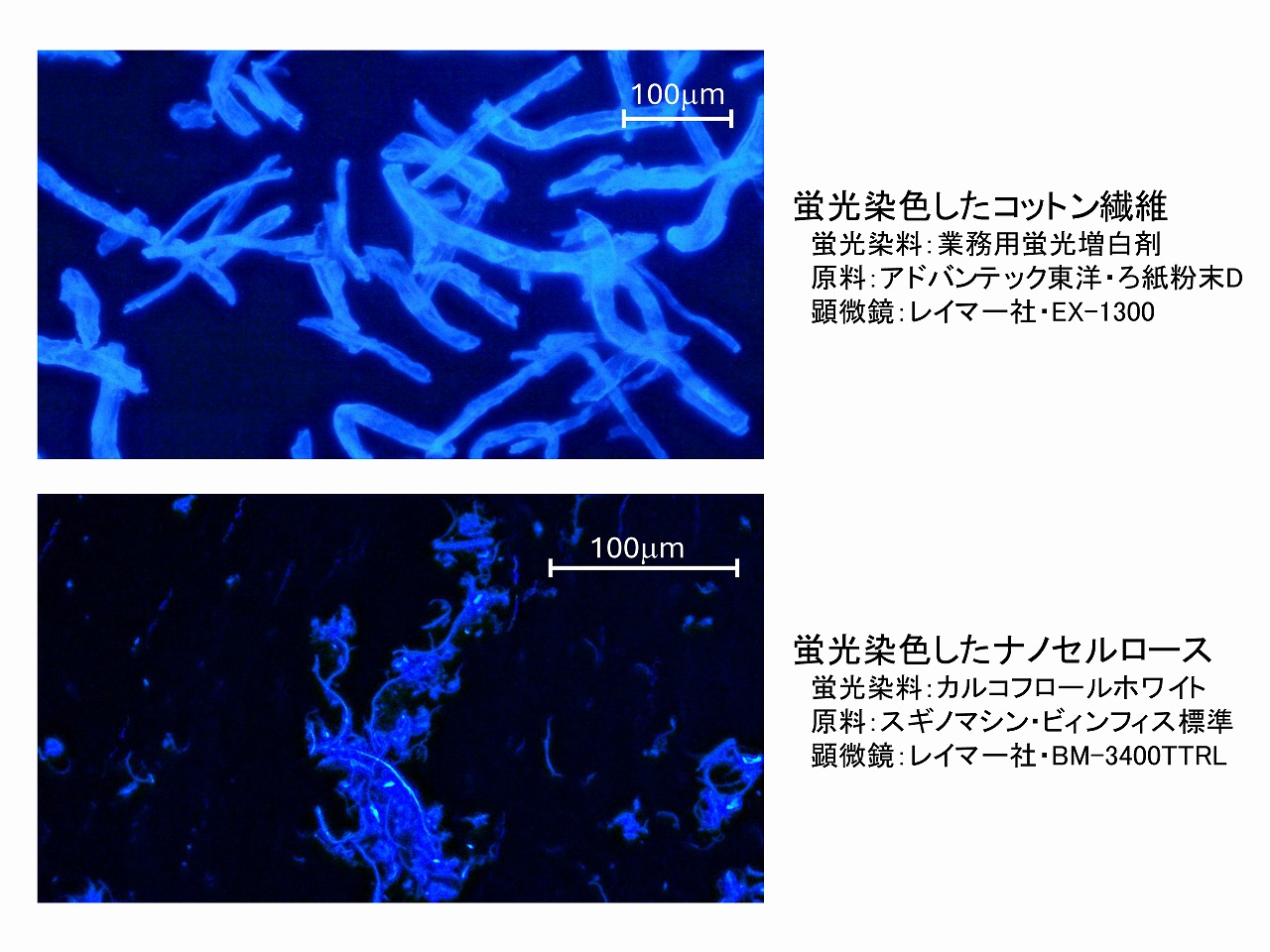
上図は、LED UVランプをプレパラートの上部横から照射して観察した、蛍光染色済みのコットン繊維とナノセルロースです。本格的な蛍光顕微鏡と比較すると解像度等は低くなりますが、とても簡単に蛍光顕微鏡画像が取得できます。
本格的な蛍光顕微鏡は、対物レンズを通してUV等をサンプルに照射するため、対物レンズの材質は、UVを吸収して発光しない材質が求められ、比較的高価です。しかし、プレパラートの上部横からLED
UVランプで照射しているため、途中にUVをを吸収して発光するものはなく、蛍光で光っているサンプルのみを顕微鏡観察しているため、顕微鏡は高価なものでなくても、それなりに観察することができます。
この方法は、上部横からUV照射してるため、対物レンズの倍率が大きいと、「作動距離」が小さくなり、対物レンズ先端とプレパラートが接近するため、照射できなくなります。低倍率観察か作動距離の比較的大きい対物レンズを用いると多少は改善できます。金属顕微鏡は、作動距離の大きい対物レンズが用いられている場合が多いです。
上図の写真は、蛍光染色したコットン繊維を比較的低コストな学習用顕微鏡で観察した場合と、蛍光染色したナノセルロースを金属顕微鏡で観察した場合の例を掲載しています。蛍光染色したセルロースの蛍光は強く、暗室などは使わず、通常の照明が付いたままの実験室で取得しています。蛍光染料は、UVで光りますが、長時間照射し続けると蛍光染料が分解して光らなくなります。
以上のような、簡易的な蛍光顕微鏡は簡単に実施できますが、紫外線の危険性や作用などは十分に理解して、各自の自己責任で実施して頂けたらと思います。
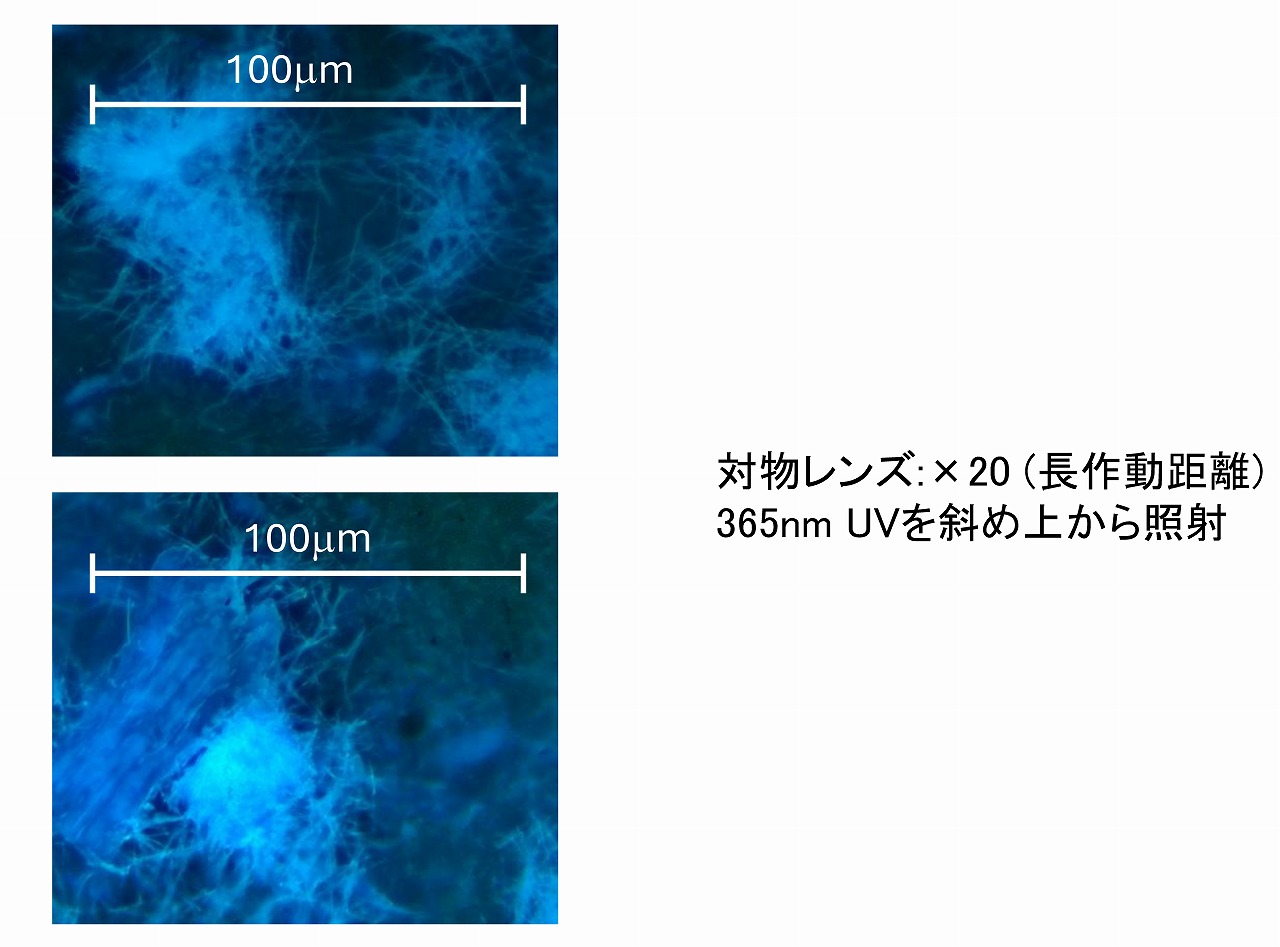
照射条件などを最適化すれば、一般的な透過型顕微鏡でも、下図のような写真は撮影することができます。ここでは、5Wの365nmUVライトで照射し、(株)レイマー・BM-3400TTRLの20倍の対物レンズで、反射モード、CCDは(株)レイマー 顕微鏡用マルチインターフェース4KカメラFLOYD-4Kを使って撮影しました。レイマー社のCCDは専用ソフトを用いることで、ゲイン・シャッター速度を調整でき、さらにZ軸スタッキングモードを活用すると画像が積算されてキレイな写真が撮れます。
コットン繊維と木材パルプ繊維の区別方法

セルロース系の素材を用いた研究開発や製品では、建材等を別にすれば、粉末状で利用する場合も多いと思います。上図は、それぞれコットン(高度に精製された綿)、精製木材パルプ、自作したスギ木粉ですが、マクロ的に観察しても色以外では区別が難しいです。特に、コットン粉末と精製木材パルプ粉末は見た目だけでは区別できません。
正しいルートで入手すれば、問題にならないかもしれませんが、何か分からないが、セルロース系の粉末がある場合、これが何なのかは知りたくなります。もし、樹脂粉末等であれば燃やしてみればその性状の変化(溶融する等)や煙・臭いから判断も出来るかもしれません。繊維分野では、燃やして臭いで判断することも行われるそうです。しかし、純度や劣化度合いなども含めて評価したい場合もあるかもしれません。
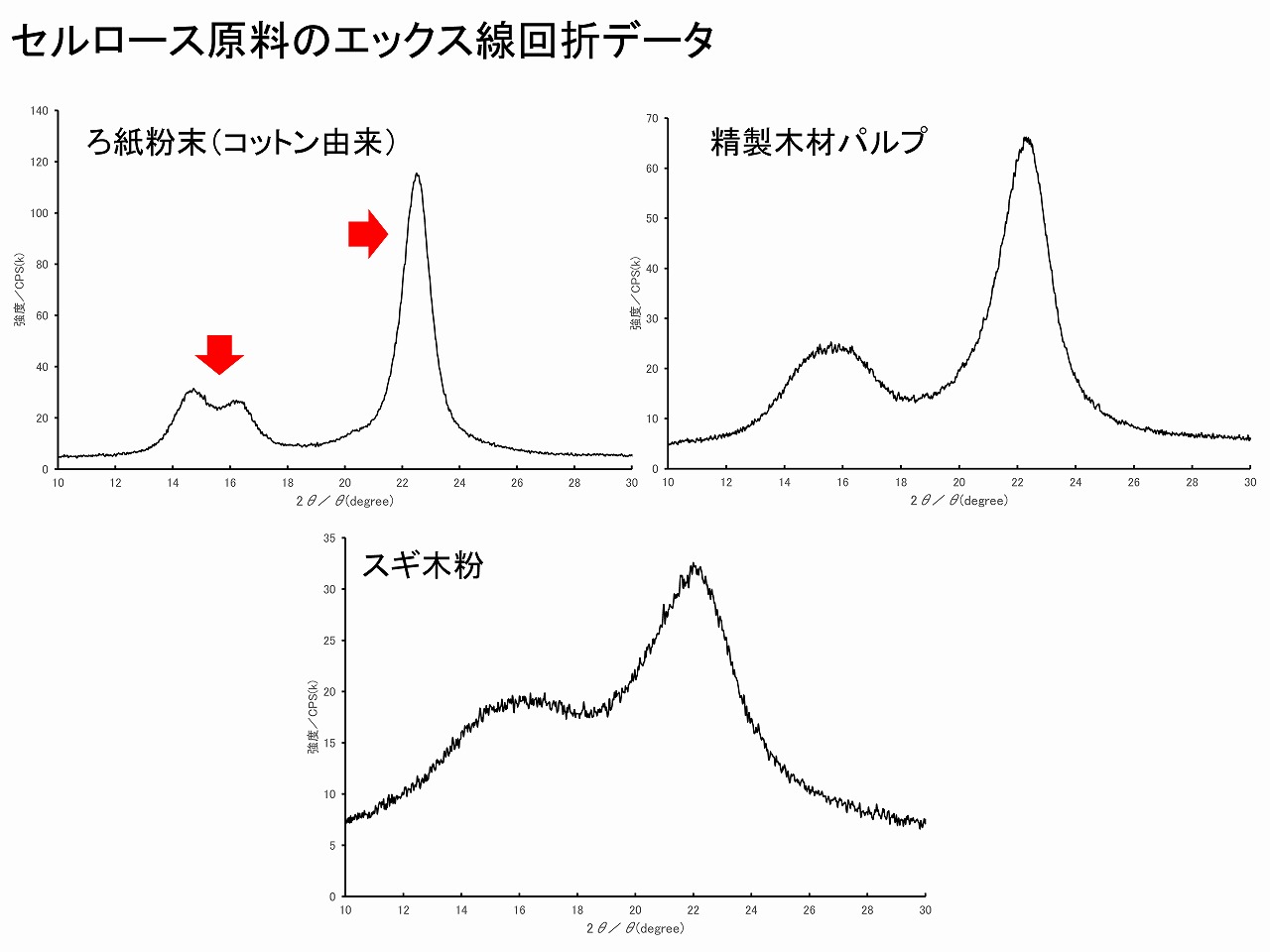
白っぽい物質がセルロースかどうかは、エック線回折を測定すると簡単に分かります。回折パターンは、本ページの「セルロースの結晶性(エックス線回折測定)」を参考にしてください。セルロースのエックス線回折パターンは、無機物等に比べて、ピークは小さいですが、そのパターンは特有であり、このパターンが観測されなかったら、別の物質です。過去に、天然系で得られた白っぽい物質がセルロースかどうか調べた場合、セルロース特有の回折パターンが見えず、他の分析からヘミセルロース系物質であることが分かったこともあります。エックス線回折は、ある意味特殊で、設置には事前に労働基準監督署への手続きが必要となりますが、測定そのものはとても簡単にできます。
上図には、それぞれコットン(高度に精製された綿)、精製木材パルプ、自作したスギ木粉のエックス線回折パターンを示していますが、コットンのみ少しパターンが異なります。精製木材パルプとスギ木粉は類似していますが、スギ木粉の方がピークがブロードに見えるのは、精製木材パルプと異なり、ヘミセルロースやリグニン等の非晶質成分が多いためです。コットンでは、非晶質成分が少ない上、セルロースの結晶子サイズが木材より大きいため、回折ピークはシャープになります。また、15度付近には2つの小さなピークも見えます。コットンの場合でも、結晶性等が低下しているとこの2つのピークはそれぞれがブロード化して1つに見えることもありますが、ピークのシャープさは木材系とは区別しやすい場合は多いです。上図のコットンの典型的パターンが見えは場合は、ほぼコットンと判断しても良いです。バクテリアセルロースやナタデココ、セロハン等は、また少し異なった波ターンを示しますので注意は必要ですが、基本的パターンは類似しています。
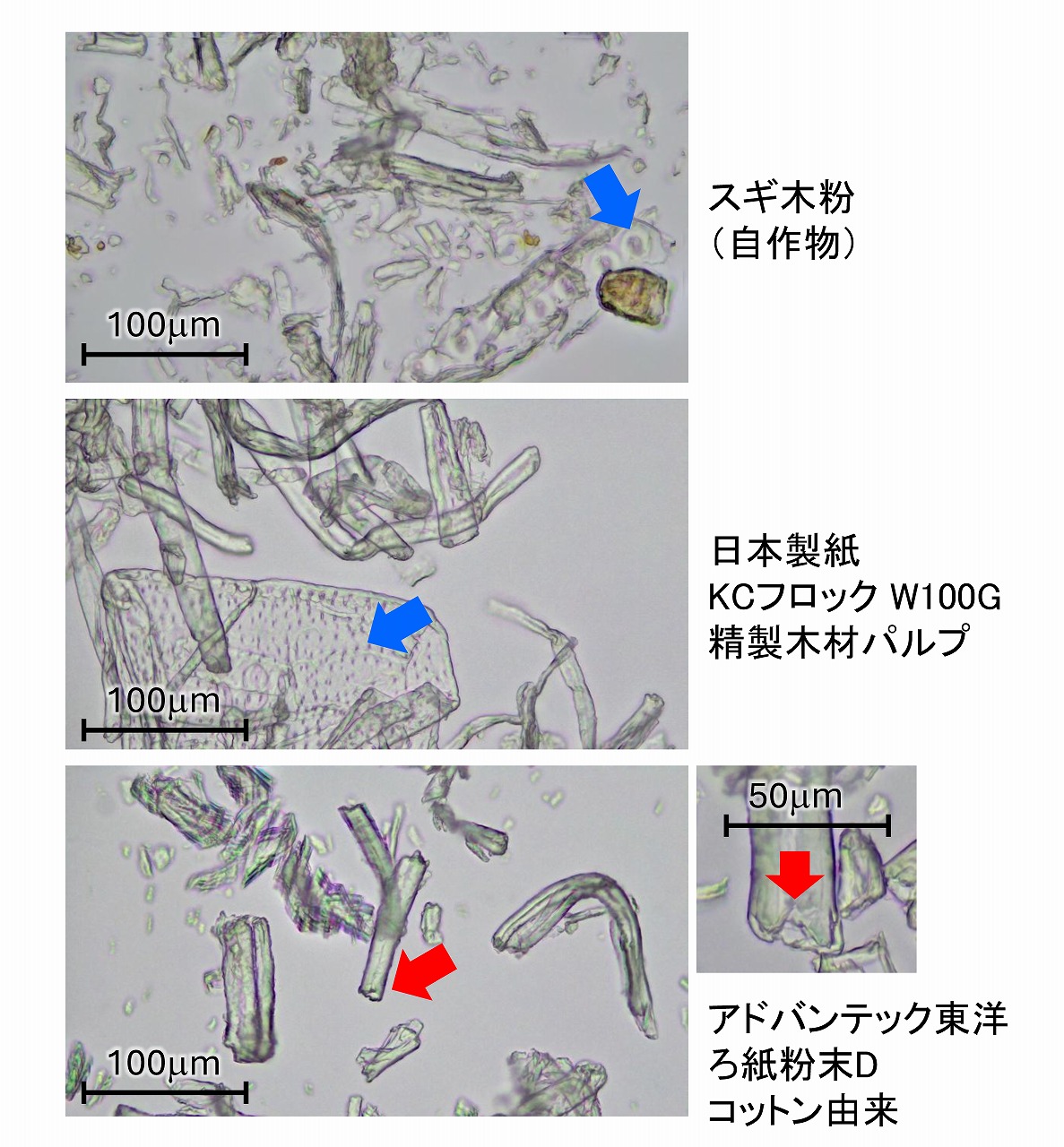
エックス線回折測定は、セルロースの結晶性の評価にも役立ちますが、簡単に使用できない環境もあります。そのような場合、コットンなのか、木材パルプなのかの判断は、普通の光学顕微鏡でも評価できる場合があります。上図は、それぞれコットン(高度に精製された綿)、精製木材パルプ、自作したスギ木粉を水に分散させてから、スライドガラスに滴下し、カバーガラスをかぶせてから光学顕微鏡で観察した事例です。
コットンと木材パルプ・木粉の区別は、その形状からも簡単に分かります。コットンは繊維サイズのバリエーションがそれほどなく、おおよそ同じサイズで形状は比較的直線的です(湾曲したものもあります)。木材パルプや木粉は、分級してあってもサイズや形状が比較的バラついている場合が多いです。また、上図中で青矢印で示した部分には、吸盤状構造やメッシュ状構造が見えます。これは木材組織に元々ある「壁孔」と呼ばれる構造で、木材組織の仮導管の壁などにあり水などの輸送に使われている構造です。上図の精製木材パルプにもメッシュ状構造がみられ、この素材が確かに木材を原料としていることが分かります。コットンには、そのような吸盤構造やメッシュ構造はありません。精密に「壁孔」の構造を調べると木の種類も同定できる可能性もあります。
上図の写真はろ紙粉末はコットン由来であり、コットンにしか観察されない構造が見えています。コットンは中空構造を持ったホローファイバーです(赤矢印部分)。中空であることが明確に分かる部分を観察するのは簡単ではないですが、よく調べると分かります。乾燥したコットン繊維は、潰れていて中空構造が観察されにくい時もあるため、水中で膨潤させてから光学顕微鏡観察するなども必要になる場合があります。中空構造が観察されたら、その素材はコットンと判断できます。微粉砕等の処理がしてなければ、精製しても中空構造は残っています。化学処理等では中空構造が消える場合もあります。
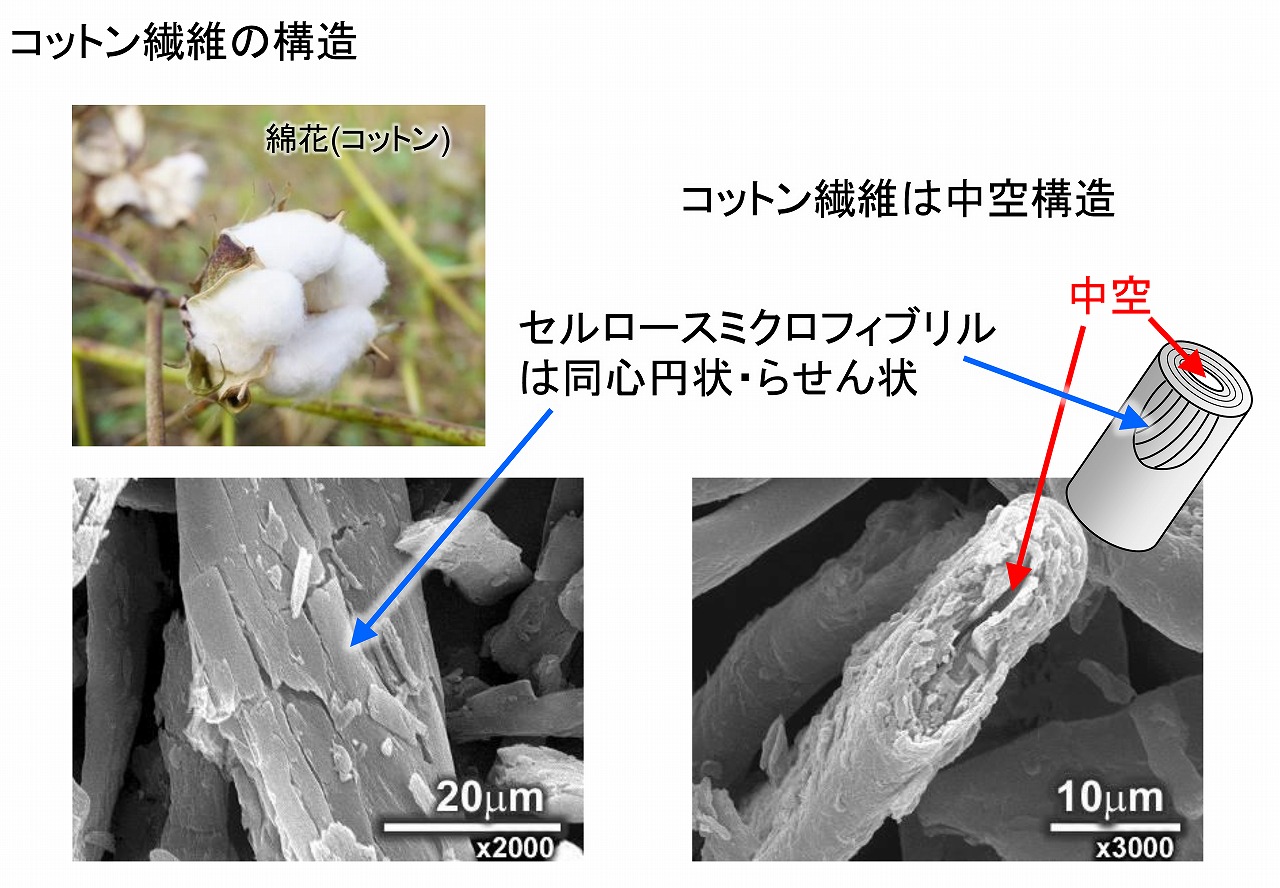
コットン繊維は、木材パルプ繊維とは明確に異なってたいます。コットン製品は多く市場にありますが、コットンの中空構造で、保温性や吸湿性などが発揮されており、特に水分を吸うと乾燥時よりも強度が向上する性質から、衣類には無くてはならない特性を持っています。
コットンは中空の中心の周りにセルロースミクロフィブリル(セルロース分子の束)が同心円状かつらせん状になっています。上図の写真は、電子顕微鏡で観察したコットン繊維です。中空構造とセルロースミクロフィブリルの積層構造が分かります。
簡易的偏光顕微鏡観察
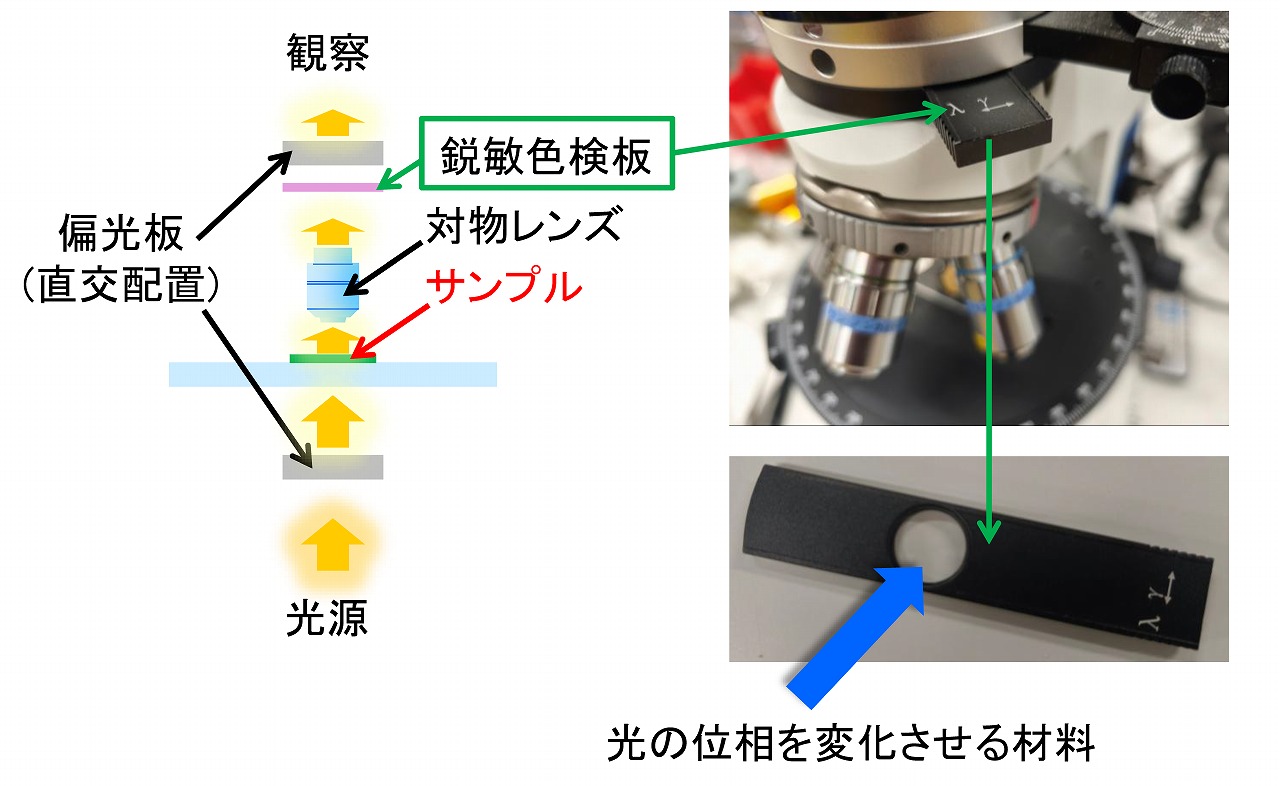
高分子材料や鉱物等の評価・解析では、偏光顕微鏡はよく使われる分析装置です。偏光顕微鏡を用いることで、高分子材料では分子の配向に関する情報が得られます。鉱物では、その構成物に関する情報が得られます。
偏光顕微鏡の基本的な応用事例は、本ページの「セルロースの偏光顕微鏡観察」、「偏光顕微鏡によるセルロース繊維の評価」をご参照下さい。
偏光顕微鏡は、基本的には、光学顕微鏡です。透過型の光学顕微鏡に、偏光板が2枚ほど取り付けられています。さらに、観察を容易にするために、鋭敏色検板も取り付けられるものもありまが、メーカーによっては、オプションになっている場合もあります。上図には、偏光顕微鏡での、偏光板と鋭敏色検板の位置関係を示しています。偏光顕微鏡は、用途によってはとても便利ですが、一般的な透過型の光学顕微鏡と比較すると高価です。一般的な透過型の光学顕微鏡は、別の言い方では、生物顕微鏡とも言われます。入門用から研究用まで様々ですが、比較的安価なものでも、信頼できるメーカーであれば、思った以上にキレイに見える場合はよくあります。光学顕微鏡の性能(見え方)の相当部分は対物レンズの性能で決まります。しかし、キレイに見えても、生物顕微鏡で高分子材料や鉱物等を見ても、偏光の情報は得られません。オプションで、生物顕微鏡を偏光顕微鏡のようにできるものもありますが、それほど、安価でもありません。簡易テストで高分子材料での分子の配向等を調べたい場合、専用の偏光顕微鏡を新たに購入するほどでもない場合、小中高などの学校での理科教育や実習などで簡単に偏光が調べたい場合、そのような場合には、既存の生物顕微鏡を用いてサンプルを偏光シートと位相差シートを使うことで、簡易的ですが、偏光顕微鏡の画像が取得できます。
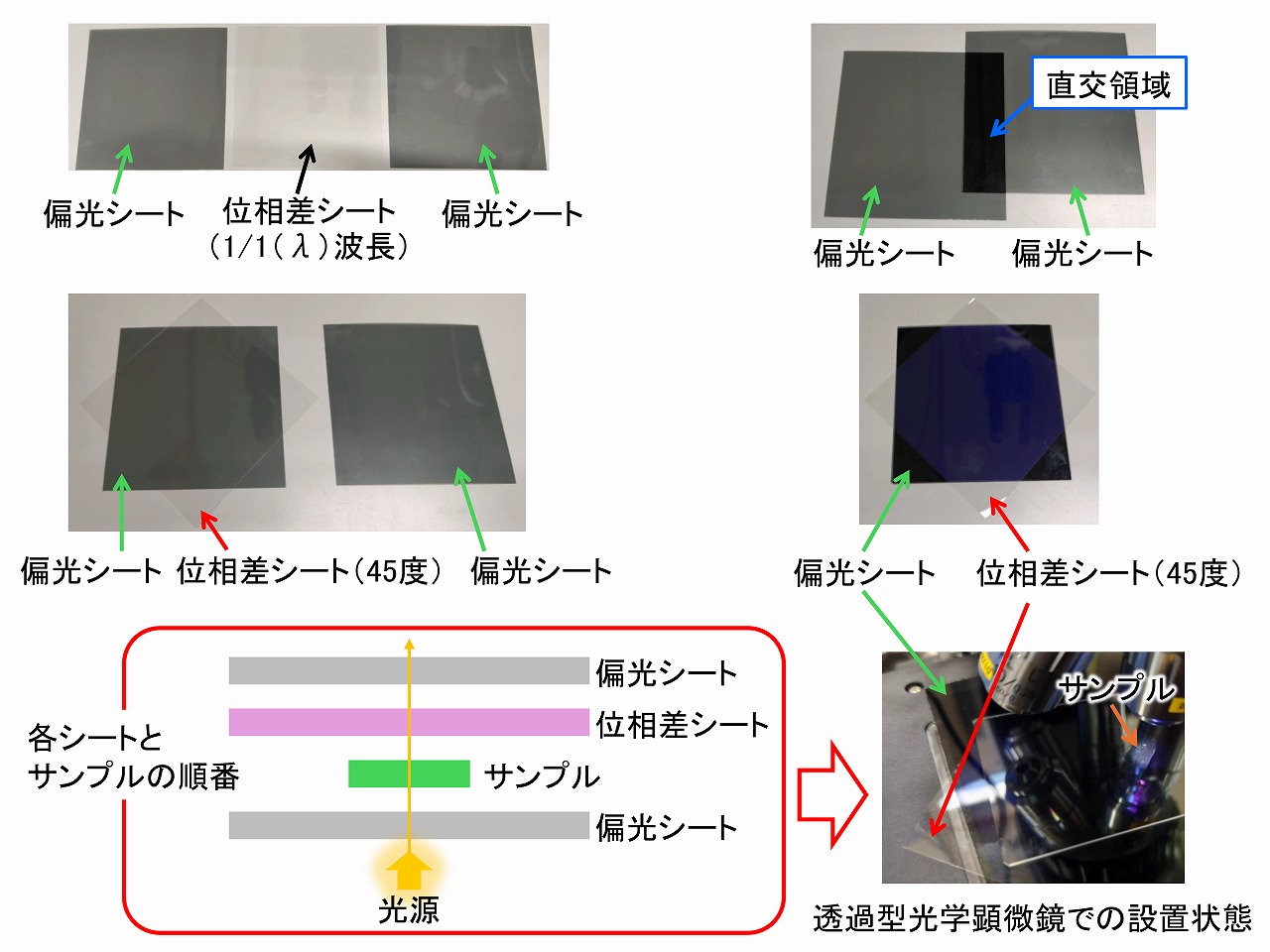
上図は、今回用いた、偏光シートと位相差シートの事例です。偏光観察では、偏光シートは2枚必要です。基本的な偏光顕微鏡観察では、「セルロースの偏光顕微鏡観察」に記載しているように、偏光シートは、直交状態で使います。位相差シートは、光の位相を変化(ずらす)させるもので、鋭敏色検板(鋭敏色板)と呼ばれています。これを用いると、偏光の色(レターデーション/干渉色)が大きく変化して、とても明確な色を持った偏光画像になります。鋭敏色検板には、1/1(λ)波長、1/4波長の他にも、いくつかあります。よく用いられるのは、1/1(λ)波長です。
偏光観察では、偏光シート、位相差シート、サンプルの順番が大切です。さらに、偏光シートと位相差シートはその向きも大切です。上述の様に、偏光シートは、直交状態とするのか基本です。直交では光が通りにくいため暗くなります(上図の直交領域)。位相差シートは、そのままでは方向が判断しにくいですが、実際にサンプルをセットしてから確認することができます。偏光シートも位相差シートもメーカーによって向き(角度)が異なる場合があります。
偏光シートは、理科実験などでもよく使われるため、それほど高価では無く、入手は容易です。しかし、位相差シートは、それほど汎用的なものではありません。今回は、以下に示したショップから入手した1波長シートを用いました。ここのショップのホームページには、偏光や蛍光、顕微鏡などに関する製品情報の他、解説も充実しています。
◇入手先
株式会社美舘イメージング、1波長位相差板/MGR570-2(500x500mm)

上図は、偏光シート・位相差シート・サンプルの有無でどのように画像が変わるかを示しています。右下の順番で観察した場合には、とても鮮やかな色が見えています。用いたサンプルは、ポリプロピレン(PP)を溶融させて、加熱プレス方法でシート化し、急激に温度を低下させないようにして、ゆっくりとエージングしたものです。見えている放射状の構造は、ポリプロピレンの球晶です。
このような、単にサンプルをシート挟んだだけの簡易的な方法でも、高分子材料の結晶を明確に観察することができます。単にシートを重ねているだけであるため、光学的な平面性は高くなく、画像がゆがんだり、ピントが正確に合わない場合もありますが、操作そのものは、とても簡単なため、簡易的チェックや現場などでの品質確認、教育現場などでは活用できるのではと考えられます。
セルロースサンプルの乾燥方法
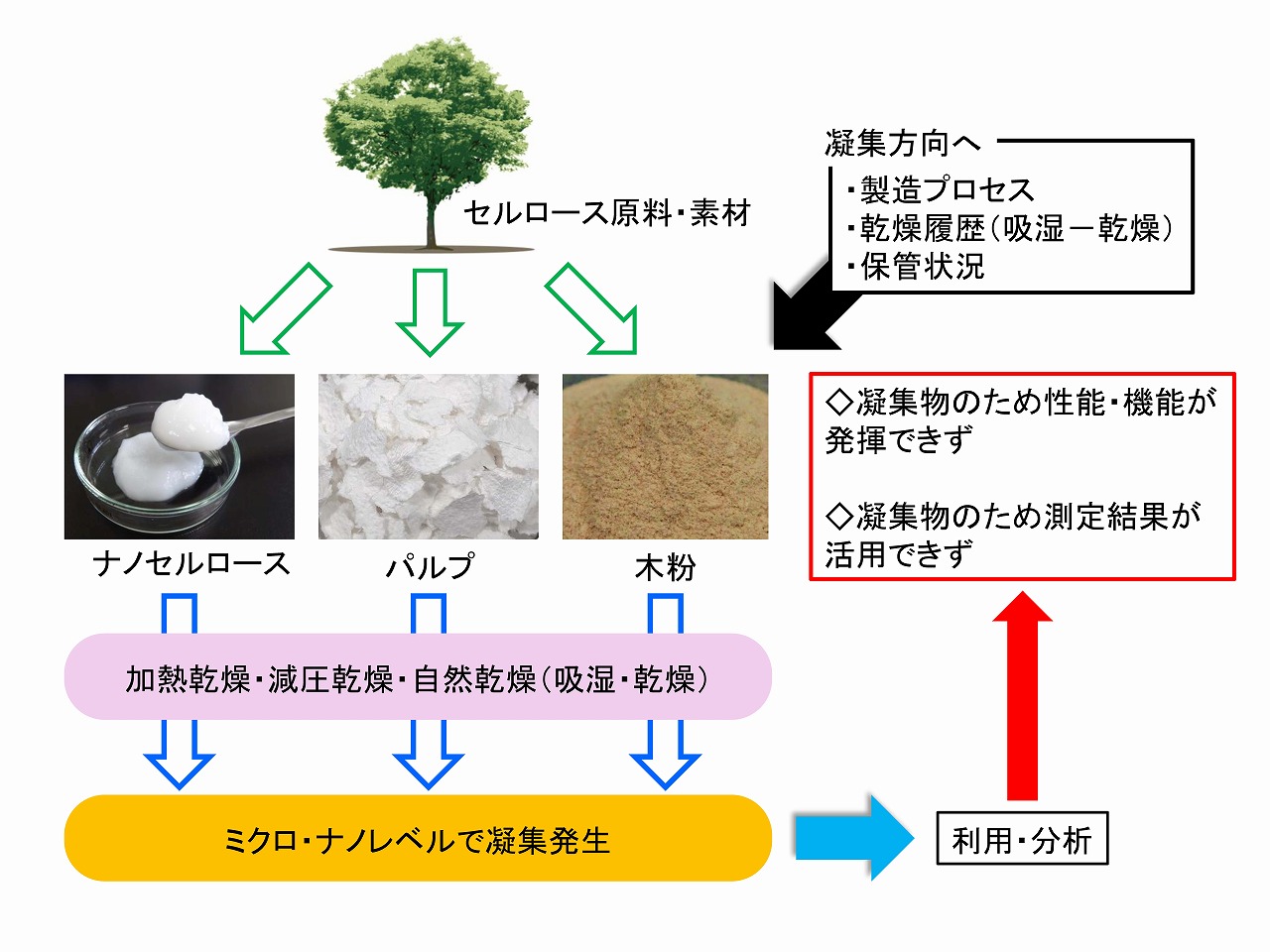
パルプやコットン、木粉等のセルロース原料を使って様々な実験を行う場合、普段、そのセルロース原料の、手元に来るまでの経過や工程などは、気にしていないことが多くあります。セルロース誘導体の研究、ナノセルロースの製造研究、添加剤等としてセルロース利用の研究などを行った場合、上手く行かなかったり、再現性が悪かったり、季節やその日の天気が結果に影響したりする場合もあります。手元に来るまでのセルロースの歴史は、「黒歴史」かもしれません。
セルロースは、水ととても馴染みが良く、普通に放置していても保管環境によって、吸着水分量は3%くらいから10%程度で変化しています。雨の日には、10%くらい吸水しており、冬の乾燥した季節では、3%程度になっていることもあります。このような、吸湿と乾燥を繰り返して行くと、セルロース原料は、ミクロ的にもマクロ的にも凝集していきます。特に、ナノのセルロースミクロフィブリルレベルで凝集すると、反応性などは低下します。また、セルロース原料の比表面積を調べようと思った場合、比表面積は小さく出ます。セルロースは、基本的に凝集する物質です。これは、例えばコットンのようなセルロース純度の高い原料から製造したナノセルロースは、低濃度に保持していても、水中で凝集する場合があります。カルボキシル基を持つTEMPO酸化ナノセルロースなどや、イオン性官能基を持つ木材から直接的に製造したナノセルロースは、水中でも静電的反発で凝集しにくいと言われています。
凝集しているのか、凝集していないのかの区別は、簡単にはできません。凝集度合いが、結果に影響すると考えられたら、その凝集の歴史をできる限りキャンセルさせる必要があります。これまでの実験等では、一度、水に浸漬(3日程度からサンプルによっては1週間)してから、凝集を抑制する方法で乾燥するというのが、有効でした。以下に、その手順の概要を示しました。
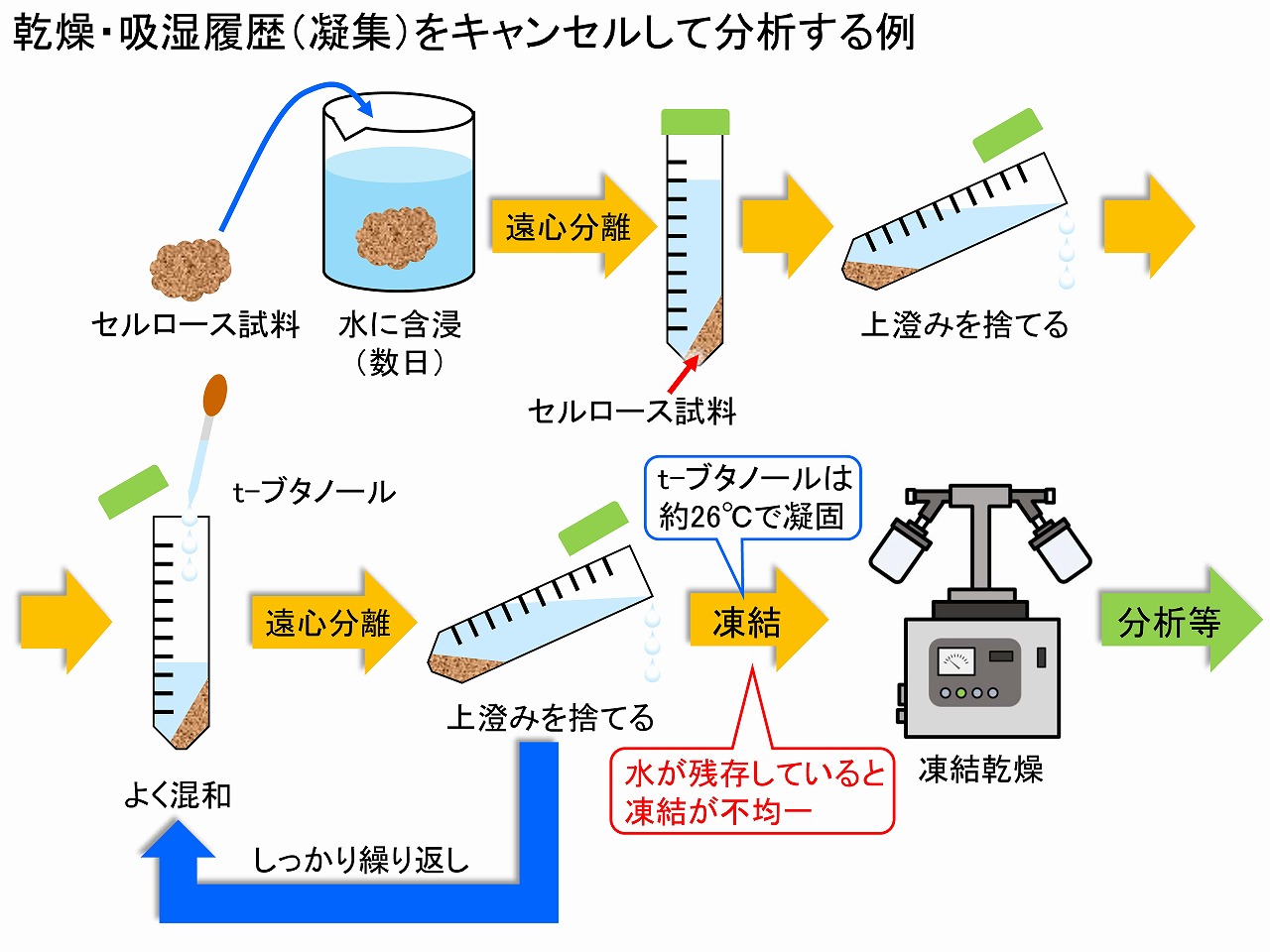
t-ブタノールを用いる方法は、電子顕微鏡用の声帯サンプルの乾燥法等で、古くから使われている方法です。理論的には、系中の水を完全にt-ブタノールに置換しなくても、凝集は押さえられますが、置換が全体として均一かどうかの判別は難しいため、単純には、できるかぎり100%t-ブタノール置換をされる場合が多いです。水が残っていると、そこは凍結乾燥時に凝集しやすくなります。この、t-ブタノール置換は単純な作業ですが、しっかりと何度(10回以上)も実施することが大切と言われています。置換作業後は、冷蔵庫で凍結の具合を見ます。t-ブタノールは約26℃で凝固しますが、水が残っていると、凍結の仕方が不均一になります。置換が確認できたら、最後に凍結乾燥器等にセットして、凝固したt-ブタノールを昇華させます。この減圧乾燥器の操作では、t-ブタノールが真空ポンプの方に来るため、トラップ温度は、-90℃程度の低温型が好ましく、真空ポンプも耐食性の高いポンプの方が推奨されています。しかし、メーカーではt-ブタノールによる劣化は保証しないこともあるようなので、高頻度での凍結乾燥器のチャンバーの製造、真空ポンプのオイル交換が、装置寿命を長くするためにも大切です。
このようにして、凍結乾燥器で乾燥処理した、セルロースサンプルは、比較的早く、乾燥状態になったように見えます。しかし、t-ブタノールは簡単には無くなってくれません。実際、我々の実験では、凍結乾燥3日目のサンプルを分光的に調べたところ、相当量のt-ブタノールが残存していました。そのため、節密分析用のサンプル調整に限らず、1週間以上、凍結乾燥器・減圧乾燥することを基本としています。このような、セルロースが有機溶媒等を保持する現象は、80年以上前から知られています。以下に、その現象を深く研究した、「ヘルマン・シュタウディンガー/1953年ノーベル化学賞受賞」の論文やノーベル賞受賞講演の翻訳本からの内容を簡単に記載しました。
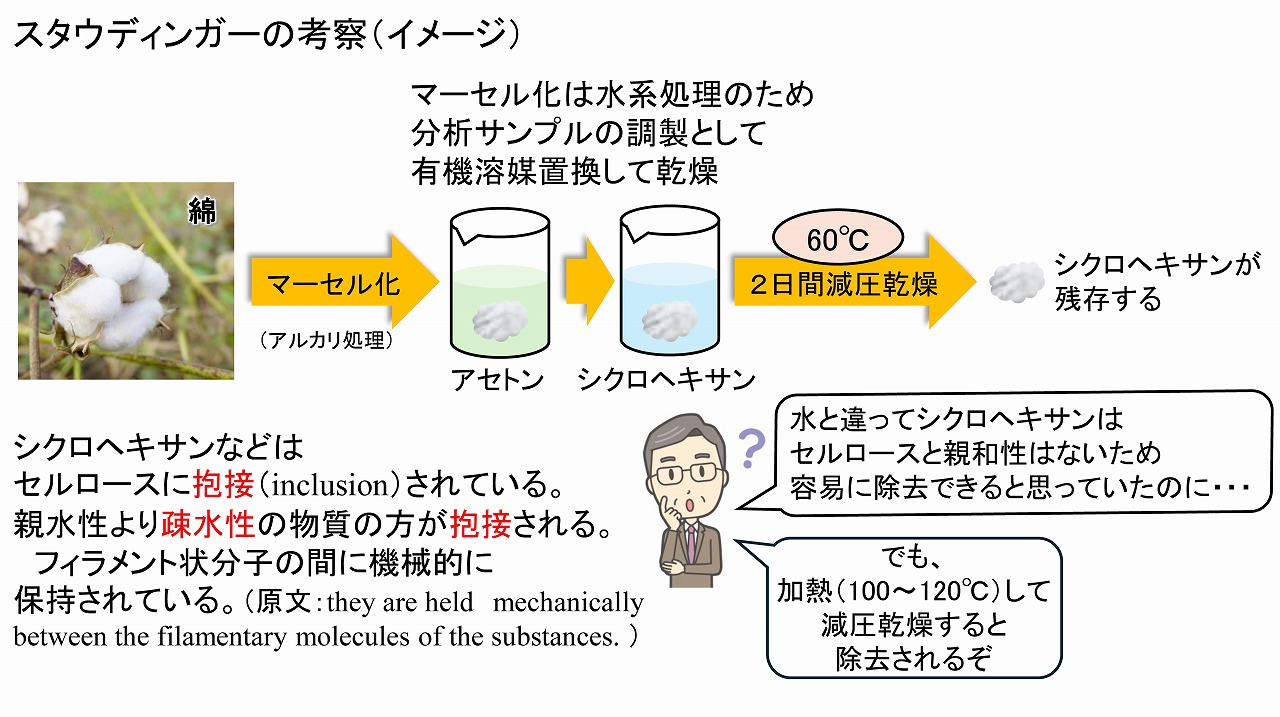
「ヘルマン・シュタウディンガー」は、ドイツの科学者です。高分子化学を幅広く研究し、1953年にノーベル化学賞を受賞しました。2025年のノーベル化学賞は、北川 進 先生で、様々な物質を選択的に吸蔵する、多孔性有機金属構造体(MOF/Metal
Organic Framework)の開発がノーベル化学賞の受賞理由となっています。セルロースも、選択制は高くはないものの、有機分子を吸蔵する性質があることは、実は、とても昔から知られていました。
シュタウディンガーに関する文献等を見ると、セルロースによる有機分子の吸蔵現象や、それによる化学的反応性の変化などを、様々な面から研究されています。特に、セルロース原料が、有機分子を吸蔵して、取り除くのが簡単ではないことが何度も示されています。つまり、上記の、t-ブタノールも、置換個-凍結乾燥後では、簡単には除去できないことが古くから知られていたと言うことです。特に、精密な分析や実験に使うためには、しっかりと減圧下で乾燥、場合によっては、加熱して減圧乾燥が必要です。
※参考1:Hermann Staudinger, Macromolecular chemistry, Nobel Lecture, December
11, 1953.
(概要版は、ノーベル賞財団のホームページで見ることができます)
※参考2:シュタウディンガー、研究回顧、高分子科学への道、小林義郎 訳、岩波書店、1966年8月30日.
(古書としてもに入手可能です。今読んでも、高分子化学のとても参考になる内容です。)

樹脂複合材での木粉の露出
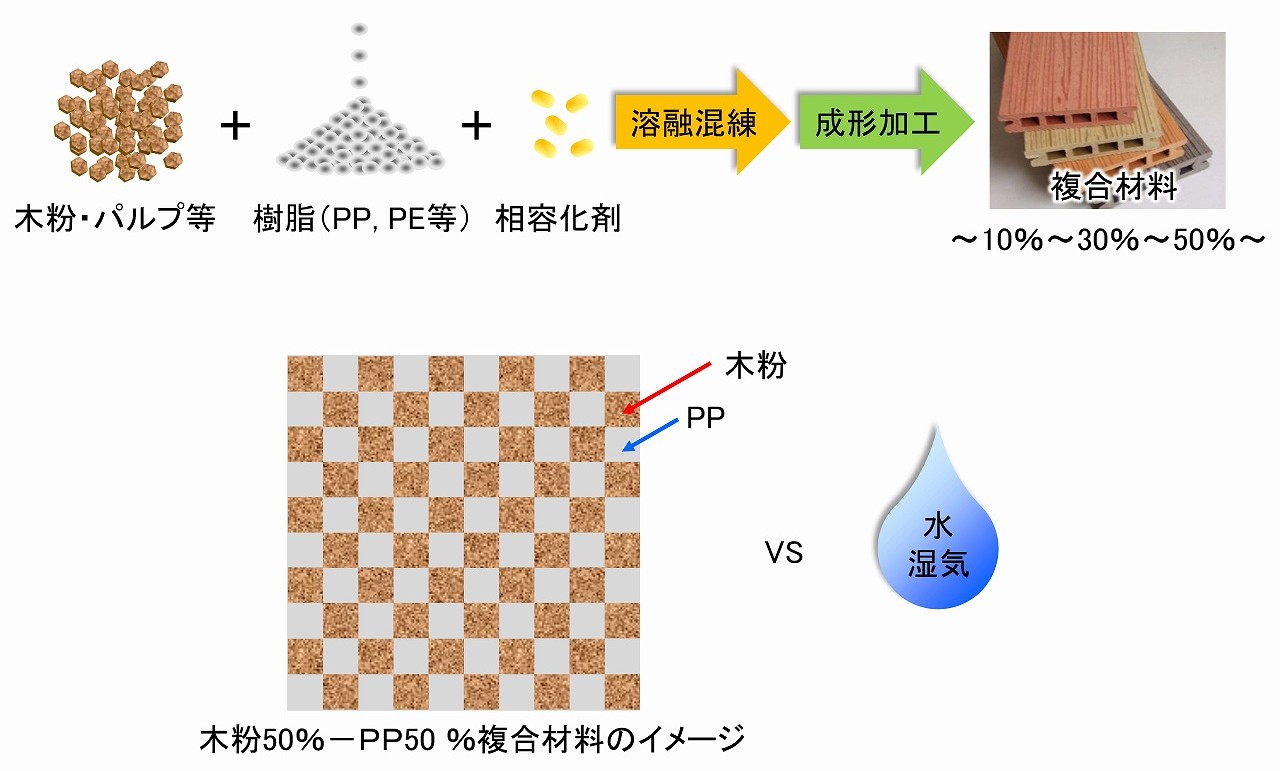
木粉やパルプなどのセルロース原料をPP等の樹脂と複合化した材料は、30年以上前から市場に出ています。木粉と樹脂の複合材は、「基礎解説/ウッドプラスチックコンポジット(WPC)の基本」にも解説しています。」このような、木粉やパルプなどのセルロース原料の複合化では、セルロース原料の複合化する割合は、10%程度から、様々な材料があります。また、性能向上目的以外にも、容器リサイクル法対策や再生資源であるセルロース原料の材料としての固定量を増やすために、50%以上も複合化される場合も多くあります。
木粉等を50%複合化した場合のイメージは、上手に乗せていますが、木粉やパルプは、水を吸収する素材なので、木粉等の複合材料は、それらが表面に露出していると、水に弱い感じがします。ナノセルロースの様な新素材を活用した材料でも、耐水性などは多くの方が気になっているようです。
ウッドプラスチックコンポジット(WPC)は、エクステリア用途も多く、屋外で雨がかかる場所でも使われていることも普通です。多くの場合、WPCの表面に、耐水性の塗装などはされていませんが、10年以上の長期間でも、大きな劣化も無く使われ続けています。全ての、セルロース系原料を複合化しても、水に対しては大丈夫と言えない場合もあり、樹脂の種類や製造プロセスも材料の特性に影響します。
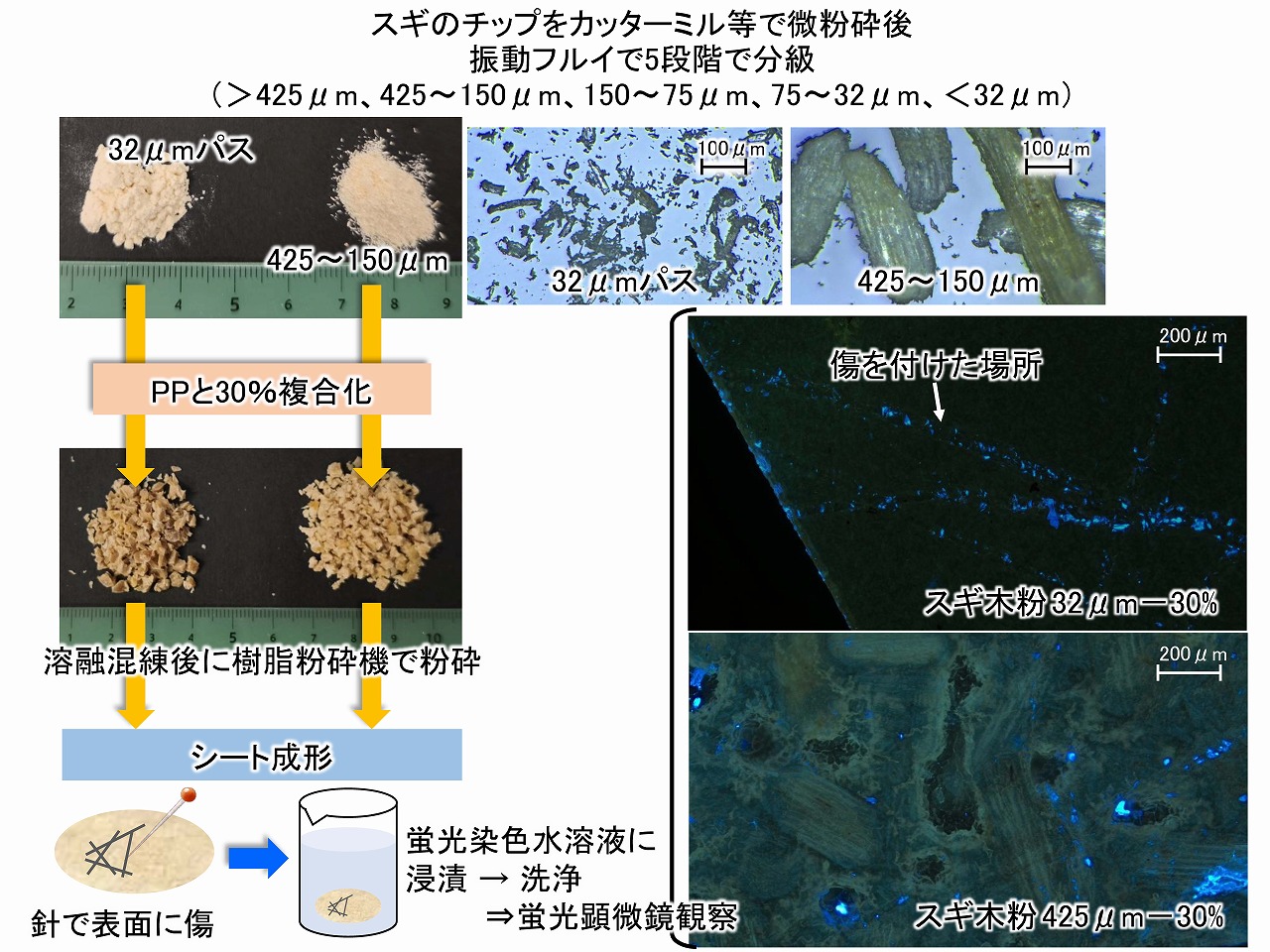
そこで、PP等に木粉やパルプを複合化した場合、どの程度の木粉やパルプが表面に露出しているのか調べてみました。下図に、スギチップからカッターミル等で粉砕して製造した木粉を、振動ふるいでふるい分けして、異なるサイズのスギ木粉を一般的な相容化剤を用いる方法でPPに複合化した実験結果を示しました。
スギ木粉は、ふるい分けで32μm以下となったスギ木粉と、425~150μmの画分のスギ木粉を、それぞれ30%でPPに複合化した材料を、簡単なプレス成形で、シート化して、シート成形体を得ました。このシート表面には、針で傷を付けました。その後、セルロースを特異的に蛍光染色する染色剤の水溶液に浸漬し、しっかりと水で洗浄してから、蛍光顕微鏡で表面観察しました。
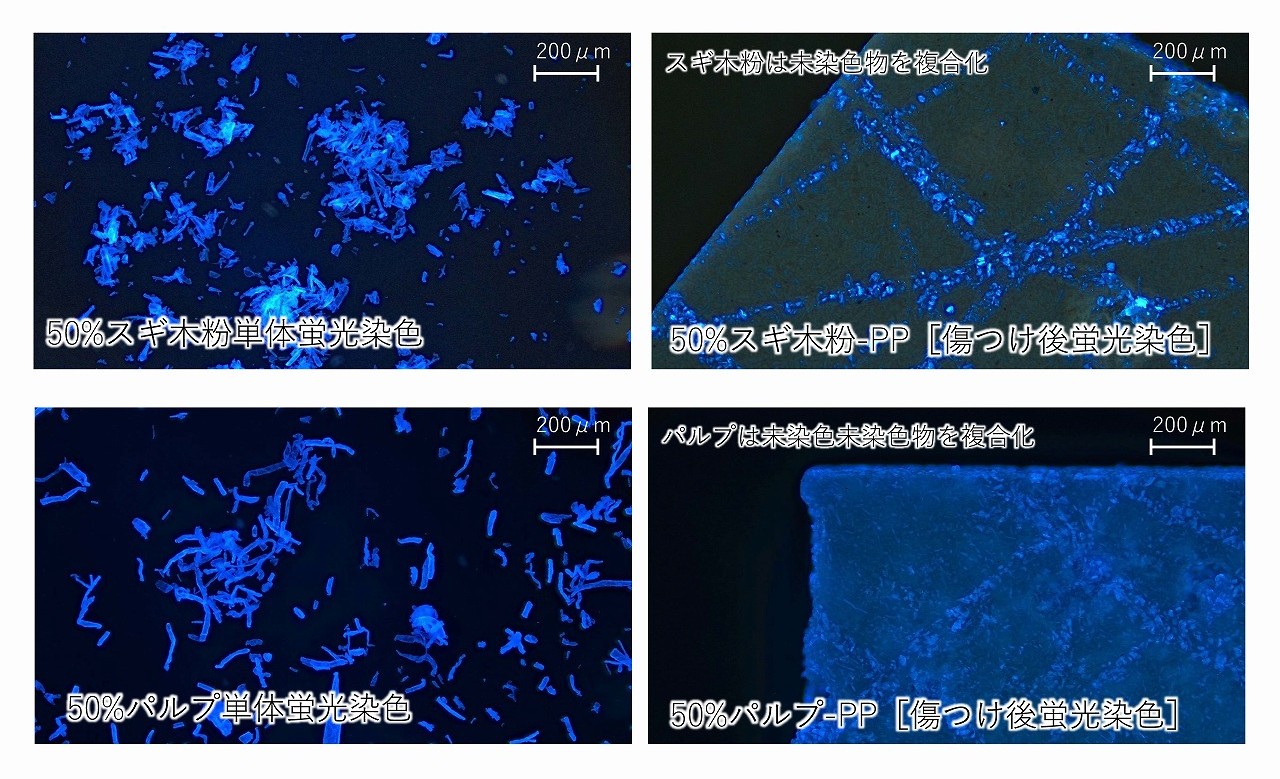
複合材料のシートの表面で、木粉が露出していれば、その木粉は蛍光染色されるはずです。実験の結果、蛍光染色されている木粉はとても少なく、針で表面に傷を付けた場所の木粉が主に、よく染色されていました。425~150μmの大きな木粉を複合化した材料では、蛍光顕微鏡の観察下で、大きな木粉の形状は確認できるものの蛍光染色されている場所は少ないことが分かりました。針で傷を付けた場所が、ほぼ特徴的に染色されていることから、複合材料の表面では、木粉はほとんど露出しておらず、表面はPP等の樹脂層(スキン層)が主であることが考えられました。傷によりスキン層が破壊されて、その層の下の木粉は染色されました。
木粉の配合量を50%に上げても、結果は同様でした。最下図では、木粉やパルプ単体は、用いた蛍光染色剤でよく染色されることを攪乱しています(左側の写真)。この場合は、32μm以下のスギ木粉を用いています。
これらの結果が示すように、セルロース系の樹脂複合材では、セルロース原料の材料・製品の表面露出は少なく、そのため耐水性も高くなることが考えられます。しかし、材料・製品の断面では、セルロース系原料は露出します。断面から、水を吸収してしまうということは、課題とされています。また、製法や原料種、成形加工条件によっては、耐水性がなくなるという話もあります。シート成形や射出成形の様な成形加工方法では、材料・製品の切断部位(スプールやランナーの切断部等)は少ないため、比較的高い耐水性が期待できます。樹脂のみのスキン層を適切に発生させるためには、成形温度、金型温度などは大切と考えられます。複合材料では、添加しているセルロース系原料は熱可塑性や熱流動性は、極めて低いため、熱流動性の高い樹脂の適切な流動性制御が、耐水性のあるセルロース系複合材料の開発では、ポイントになると思われます。
PPシート成形時の剥離材の影響
本講座では、ポリプロピレン等とセルロース系素材の複合材料に関する研究開発を進めています。このような複合材料の特性を調べる場合は、ポットプレス等を用いてシート状に成形して、切り抜いたりして各種分析装置にセットします。このような樹脂材料のホットプレス等でのシート化では、ステンレス板に挟んプレスしますが、後から成形したシートを外しやすくするためと、ゴミ等の付着を防ぐために、剥離用のシートを用いる場合が多いです。
ホットプレス加熱板-ステンレス板-剥離シート-[サンプル]-剥離シート-ステンレス板-ホットプレス加熱板、の順番になりますが、この時の剥離シートには、何が用いられているでしょうか?以前は、PTFE(商品名例:テフロン)がよく用いられていました。ただ、PTFEシートは柔らかいため、へこみや傷ができやすく、そのまま使用すると、成形したサンプルシートに、へこみや傷が転写されやすい課題があります。へこみや傷が転写されると、そのまま強度試験などに用いると、へこみや傷の部分から、破断しやすくなり、再現性が低下することも起きます。そのため、近年は、ポリイミドフィルム(商品名例:カプトン)を使われる場合が多いです。このフィルムは、硬く傷やへこみができにくく、近年は入手も容易となっています。それでも、剥離性はPTFEシートの方が高く、便利なため使われ続けています。
本講座でも、いくつかのポリプロピレン(PP)系材料を分析するために、シート化した際、ポリイミドシートでは、どうしても剥がれにくいサンプルがあり、PTFEを使ってみました。サンプルシートのエックス線回折測定(一般的な汎用広角粉末X線回折測定)を行った結果、回折パターンが、これまでのポリイミドフィルムを剥離材としても用いて作製したサンプルシートと、大きく異なっていました。本講座の客員教授である高分子結晶で著名な戸田昭彦先生にお聞きしたところ、PP等の場合、PTFEでは特別な結晶となることは、よく知られているとのご意見を頂きました。
しかし、専任教員の知人等では、PPとPTFEで起きる現象について知らない人も多くいました。そこで、汎用的なポリプロピレン(PP/ホモタイプ)のみを用いて、剥離材を変えてい、シート化し、エックス線回折、偏光顕微鏡観察を行いました。また、成形シートについては、ウルトラミクロトームで、断面切片も作製して、偏光顕微鏡観察を行いました。
◇シート作製条件
○PP・・・日本ポリプロ(株)MA3
○シート化温度・・・200℃
○エージング温度・・・130℃
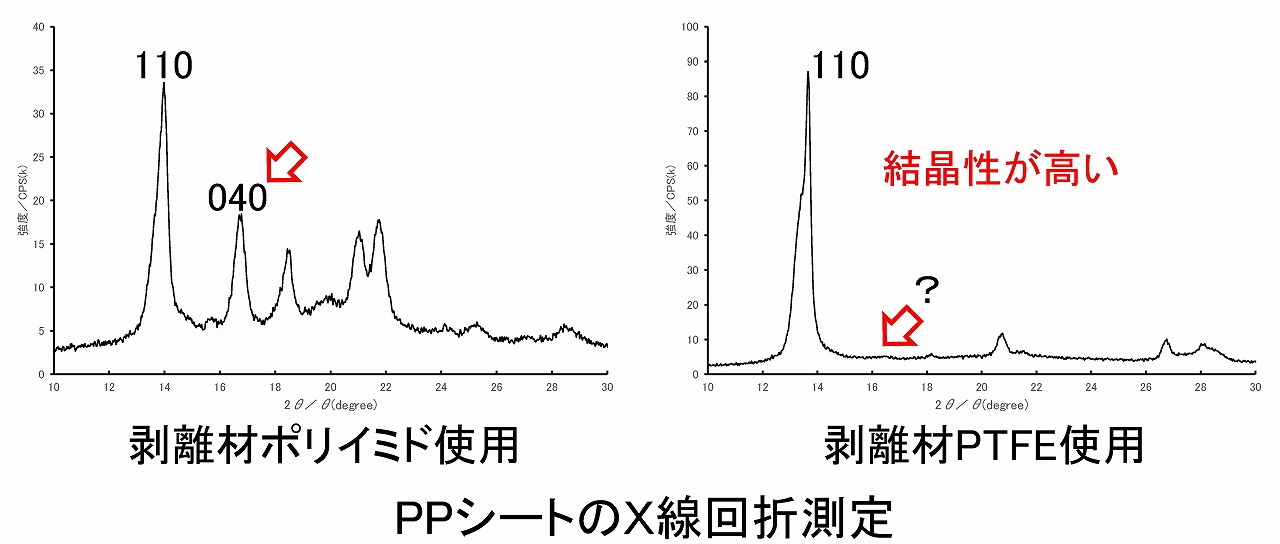
上図は、剥離材としてポリイミドフィルムとPTFEシートを使ったときのポリプロピレンシートのエックス線回折パターンです。ポリイミド剥離材では、典型的なポリプロピレンの110面と040面の回折が見えていますが、PTFE剥離材では、ポリプロピレンの110面の回折ピークが特に大きくなっています。この現象は、PTFEの表面自由エネルギーに合うポリプロピレン結晶面が優位に向くためと言われています

上図は、剥離材としてポリイミドフィルムとPTFEシートで作製したポリプロピレンサンプルシートの透過モードでの偏光顕微鏡像です。剥離材が異なるとエックス線回折パターンは、大きく異なりましたが、ポリプロピレンを溶融させてシート化し、エージングで結晶化させたサンプルシートでは、偏光顕微鏡像も、大きく異なっています。ポリイミド剥離材の場合は、透過モードで観察した偏光顕微鏡像は、典型的な大きな球晶の成長が確認されます。しかし、PTFE剥離材では、大きな球晶は確認されません。
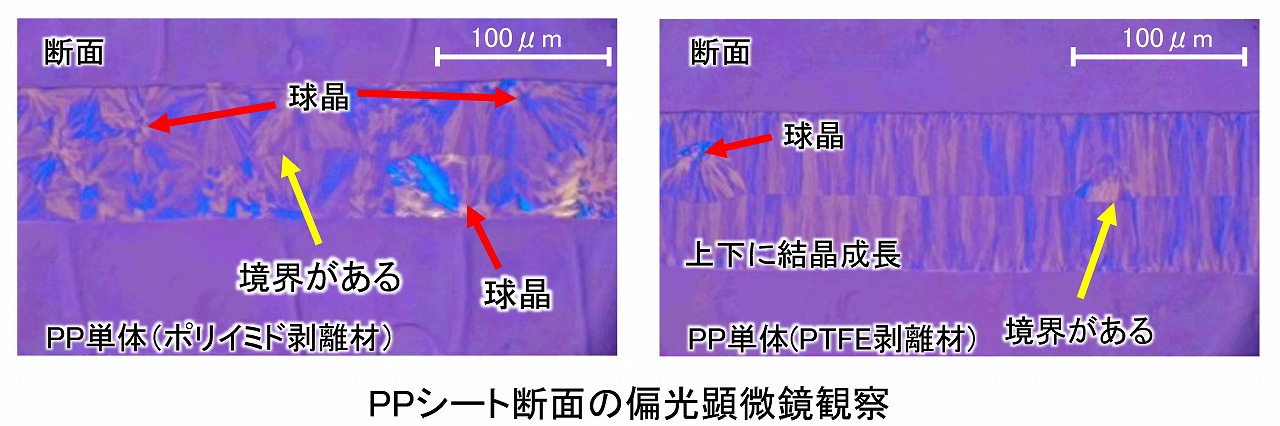
上図は、剥離材としてポリイミドフィルムとPTFEシートで作製したポリプロピレンサンプルシートを、クライオウルトラミクロトームで断面切片サンプルを作製して、偏光顕微鏡観察した結果です。
ポリイミドフィルムを剥離材としたサンプルシートでは、断面観察でも球晶が見えます。図中に「境界がある」と記載しているのは、エージング時に上下同じ温度でエージングしたことで、両面から結晶化が進行して、中央でとまったことによって境界が見えたものと考えています。
一方、PTFEを剥離材とした場合、一部、球晶も観察されましたが、PTFEの接触面から結晶が成長していることが見えました。
◎ポリイミドフィルムとPTFEシートは、材料のシート化などでは、よく使われますが、上記のように、ポリプロピレンなどでは、結晶化挙動が大きく異なっています。そのため、剥離材を適当に使っていると、結果の整合性や再現性が取れなくなることも考えられます。本講座では、ポリイミドフィルム以外に、耐熱性ポリマーシート、鉄、アルミ板等も色々とテストしましたが、結果はPTFEの場合のみ特別で、その他は類似の結果でした。PTFEによるポリプロピレンの結晶化挙動は、その材料の物性にも影響することが考えられます。PTFEは剥離性も高く便利ですが、以上のような結晶化挙動への影響もあるため、結果の解析・評価では注意が必要です。心配なときは、ポリイミドフィルムを用いる方が無難と感じています。
※上記の内容は、「第76回日本木材学会大会・広島大会」にて、ポスター発表したものの一部です。
タイトル:「I17-P1-13」「木粉系複合材料における樹脂結晶特性」
○○○○
○○○○○ ○○○○○ ○○○○○ ○○○○○ ○○○○○ ○○○○○ ○○○○○ ○○○○○ ○○○○○ ○○○○○ ○○○○○ ○○○○○ ○○○○○ ○○○○○ ○○○○○ ○○○○○ ○○○○○ ○○○○○ ○○○○○ ○○○○○ ○○○○○
○○○○
○○○○○ ○○○○○ ○○○○○ ○○○○○ ○○○○○ ○○○○○ ○○○○○ ○○○○○ ○○○○○ ○○○○○ ○○○○○ ○○○○○ ○○○○○ ○○○○○ ○○○○○ ○○○○○ ○○○○○ ○○○○○ ○○○○○ ○○○○○ ○○○○○
○○○○
○○○○○ ○○○○○ ○○○○○ ○○○○○ ○○○○○ ○○○○○ ○○○○○ ○○○○○ ○○○○○ ○○○○○ ○○○○○ ○○○○○ ○○○○○ ○○○○○ ○○○○○ ○○○○○ ○○○○○ ○○○○○ ○○○○○ ○○○○○ ○○○○○